Fotosintesi
Gli antichi Greci credevano che fosse il suolo a soddisfare tutte le
esigenze della pianta e per quasi 2000 anni questa convinzione venne da tutti
accettata come un dato di fatto. I primi successi relativi allo studio della
fotosintesi vennero accreditati al medico e chimico van Helmont nel XVII
secolo. Volendo accertare se le piante assumessero veramente tutto il loro
nutrimento dal suolo egli fece crescere in un vaso un piccolo salice non
aggiungendo al suolo nient'altro che un po' d'acqua. Cinque anni più
tardi egli calcolò che la pianta aveva raggiunto un peso di 75 Kg mentre
il suolo aveva perso solo 60 g. concluse quindi che una pianta non ricavava la
maggior parte delle sue sostanze dal suolo ma tuttavia commise l'errore di
pensare che gran parte delle sostanze presenti nel salice era stata ottenuta
dall'acqua. Un chimico inglese, Priestley, scoprì che una candela che
normalmente si spegne in un contenitore chiuso poteva continuare a bruciale
quando nel contenitore era posta una pianta e così concluse che la
pianta rinnovava l'ossigeno consumato dalla candela. Fu il medico Ingenhousz a
scoprire che le piante hanno bisogno di luce e dimostrò che esse possono
rinnovare l'ossigeno solo quando sono esposte alla luce. Solo nel XX secolo si
riuscì a descrivere questo processo nei dettagli. Per sintetizzare
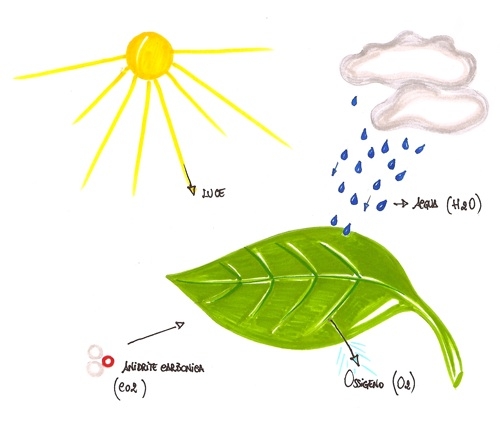
molecole di glucosio le piante hanno bisogno solo dell'energia della luce
solare, dell'anidride carbonica presente nell'aria e dell'acqua assorbita dal
suolo. L'altro importante prodotto della fotosintesi è l'ossigeno. Le
piante sono organismi autotrofi in quanto producono da sé le proprie sostanze e
non hanno quindi la necessità di nutrirsi di altri organismi o di
assumere molecole organiche. Tutti gli organismi che utilizzano l'energia
luminosa per formare molecole alimentari sono chiamati autotrofi fotosintetici.
Sulla terra ferma i principali organismi fotosintetici sono le piante,
nell'acqua sono le alghe e i batteri fotosintetici. Esistono anche autotrofi
che non sono in grado di svolgere la fotosintesi ma che si producono il
nutrimento usando l'energia contenuta in molecole presenti nell'ambiente
(chemiosintesi) tali organismi sono detti autotrofi chemiosintetici. La
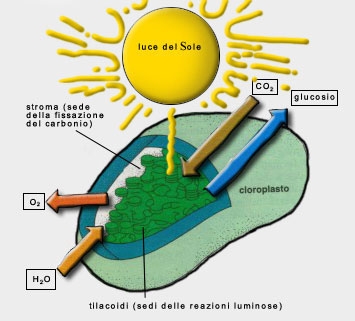
fotosintesi avviene in particolari organuli cellulari detti cloroplasti. I
cloroplasti sono concentrati nelle cellule del mesofillo, il tessuto verde
presente all'interno della foglia. Il mesofillo è costituito da due
diversi tipi di tessuto; quello a contatto con l'epidermide superiore è
chiamato tessuto a palizzata e sono queste le cellule più
fotosintetiche; il tessuto a contatto con la parte inferiore della foglia
è detto tessuto lacunoso e le cellule hanno ampi spazi tra loro che
servono a consentire la circolazione dell'aria; lo spazio tra i due tessuti
è percorso da canali che portano
alla foglia acqua e Sali minerali e che formano e nervature della foglia.
L'anidride carbonica può entrare e l'ossigeno può uscire grazie a
piccolissimi pori detti stomi, uniti a due a due e chiamate anche cellule di
guardia. Le membrane del cloroplasto formano la rete strutturale in cui
avvengono molte reazioni fotosintetiche; i cloroplasti hanno una membrana
esterna e una interna, con uno spazio compreso tra esse. La membrana interna
dei cloroplasti racchiude un secondo timento contenente un liquido denso,
lo stroma, nel quale vengono sintetizzati gli zuccheri a partire dall'anidride
carbonica. Nello stroma è immerso un complesso sistema di dischetti
provvisti di membrana, i tilacoidi, che racchiudono il terzo timento del
cloroplasto. I tilacoidi sono allineati in pile dette grana. Nelle membrane dei
tilacoidi sono inserite le molecole che catturano l'energia luminosa. Se si
osservano le piante acquatiche le foglie di queste sono spesso coperte da
bollicine. Nel XVIII secolo si ipotizzò che le piante producessero
ossigeno prelevandolo dalla molecola di anidride carbonica. Verso la
metà del nostro secolo gli scienziati hanno verificato l'attendibilità
di questa ipotesi usando un isotopo dell'ossigeno per seguire i percorsi
dell'atomo di ossigeno durante la fotosintesi. Nel primo esperimento una
pianta, cui è fornita anidride carbonica contenente l'isotopo
dell'ossigeno produce ossigeno gassoso mancante di questo isotopo (non
marcato). Nel secondo esperimento viene fornita alla pianta acqua contenente
l'isotopo dell'ossigeno e questa produce ossigeno gassoso marcato (con
l'isotopo). L'ossigeno prodotto dalla fotosintesi proviene quindi dalla molecola
d'acqua e non dall'anidride carbonica. la fotosintesi è un processo
REDOX come quello della respirazione. Quando le molecole d'acqua si scindono
liberando ossigeno si ossidano e quindi perdono elettroni insieme ad H+
. nel frattempo l'anidride carbonica si riduce a zucchero a mano a mano che gli
elettroni e gli H+ si legano ad essa. A mano a mano che l'acqua si ossida e
l'anidride carbonica si riduce gli elettroni si caricano di energia e vengono
spinti ad un livello energetico superiore; la spinta per giungere a tale
livello viene fornita dall'energia luminosa catturata dalle molecole di
clorofilla contenute nel cloroplasto.
Le tappe
della fotosintesi: La
fotosintesi non è un processo unico ma presenta due stadi ciascuno
composto da diverse tappe. Le tappe del primo stadio sono note come reazioni
luminose; questo tipo di reazioni trasforma l'energia luminosa in energia
chimica e produce ossigeno gassoso come prodotto di rifiuto. Le tappe del
secondo stadio sono note come fase oscura o ciclo di Calvin; tale processo si
svolge mediante una serie ciclica di reazioni che assemblano molecole di
zucchero a partire dalla CO2 e dai prodotti energetici delle
reazioni luminose. Le reazioni luminose della fotosintesi avvengono nelle
membrane dei tilacoidi che formano i grana del cloroplasto. La luve assorbita
dalla clorofilla presente nelle membrane dei tilacoidi fornisce energia che
aziona il dispositivo fotosintetico di formazione delle molecole organiche.
L'energia luminosa è usata sia per formare ATP a partire da ADP sia per
consentire il trasferimento degli elettroni dall'acqua al NADP+, un
trasportatore di idrogeno simile al NAD+. Gli enzimi riducono il
NADP+ in NADPH mediante l'aggiunta di una coppia di elettroni e di
uno ione H+. a mano a mano che il NADP+ si riduce l'acqua
si ossida eliminando O2. le reazioni luminose della fotosintesi sono
le tappe che assorbono energia solare e la trasformano in energia chimica
immagazzinata in molecole di ATP e NADPH. Lo zucchero si forma solo al termine
del Ciclo di Calvin. Questo ciclo ha luogo nello stroma dei cloroplasti. Il
processo mediante cui gi atomi di carbonio della CO2 sono incorporati nei
composti organici è detto fissazione del carbonio. È il NADPH
prodotto dalle reazioni luminose a fornire gli elettroni ricchi di energia
necessari ai processi di riduzione che si verificano nella fase oscura, mentre
L'ATP proveniente dalle reazioni luminose fornisce l'energia chimica necessaria
ad azionare la fase oscura. Il ciclo di Calvin non richiede direttamente la
luce.
Fase
luminosa: La luce solare
entra integra nel cloroplasto, quando esce è stata privata di alcune
lunghezze d'onda che sono state assorbite. Sulla membrana dei tilacoidi ci sono
gruppi di molecole di pigmenti (carotenoidi, clorofilla a e clorofilla b), i
fotosistemi, che hanno il compito di assorbire determinate lunghezze d'onda
senza assorbire quelle verdi. Le lunghezze d'onda assorbite sono energetiche e
questo scatena le reazioni di questa fase. Tutte le molecole dei pigmenti
assorbono energia che viene poi trasferita al centro di reazione che è
rappresentato sempre da una molecola di clorofilla a. Al centro di questa
molecola c'è un atomo di magnesio e l'energia colpisce soprattutto
questo atomo. Questa energia fa saltare i suoi due elettroni del livello energetico
più esterno ad un livello energetico superiore e subito dopo questi due
elettroni ridiscendono giù. Quando scendono si libera calore e
fluorescenza. Tutto ciò però avviene solo se la molecola di
clorofilla è fuori dal tilacoide. Quando è nella foglia i due
elettroni saltano di livello ma non scendono più perché nella membrana
del tilacoide ci sono degli accettori di elettroni. Questi accettori fanno
scendere gradualmente di livello i due elettroni facendo cedere gradualmente
energia. Ogni unità funzionale della membrana del tilacoide contiene due
fotosistemi; il fotosistema I assorbe energia fino a 700nm, il fotosistema II
assorbe energia fino a 680nm. La luce colpisce contemporaneamente i due fotosistemi; quando sono colpite le
molecole dei pigmenti assorbono energia che trasferiscono alla clorofilla a; in
tutti e due i casi l'energia quando arriva alla clorofilla a serve a far
saltare i due elettroni del magnesio ad un livello superiore. Trovano 4
accettori di elettroni situati a livelli diversi e mentre scendono liberano
gradualmente energia che serve a produrre ATP tramite chemiosmosi. Quando gli
elettroni arrivano all'ultimo accettore vanno nella clorofilla a dell'altro
fotosistema dove contemporaneamente altri due elettroni discendono lungo due
accettori di elettroni producendo NADPH. La clorofilla del secondo fotosistema
riceve i due elettroni dalla clorofilla del primo fotosistema mentre alla
clorofilla del primo fotosistema vengono ridati i due elettroni che ha ceduto
all'altro fotosistema tramite l'acqua. Una molecola di acqua si scinde in O che
si libera e 2H che danno i due elettroni alla clorofilla a liberando 2H+
(meccanismo della fotolisi). I 4 accettori di elettroni situati tra i due
fotosistemi si chiamano nell'ordine: feofitina, plastochinone, complesso dei
citocromi, plastocianina; gli ultimi due si chiamano entrambi ferredoxina
perché sono molecole della stessa sostanza anche se non perfettamente
identiche. Ogni unità funzionale si apre con il fotosistema II e si
chiude con l'ATP sintetasi. Quando la luce colpisce il fotosistema II questo
cede i due elettroni al primo trasportatore che gli è direttamente
legato (feofitina), dalla feofitina passano al plastochinone e da qui passano
al complesso dei citocromi all'interno del quale c'è un canale che
consente il passaggio di ioni H+ dallo stroma all'interno del
tilacoide. Dai citocromi gli elettroni passano alla plastocianina e reintegrano
quelli persi dall'altro fotosistema. Gli ioni H+ per gradiente di
concentrazione vanno nello stroma passando attraverso l'ATP sintetasi e
portando con loro l'energia di cui sono carichi. Con questa energia si produce
ATP a partire da ADP. Contemporaneamente a tutto questo il fotosistema I ha
ceduto i suoi due elettroni alla ferredoxina e tramite il NADP reduttasi si
produce NADPH + H+. sia ATP che NADPH servono per produrre glucosio.
La fase luminosa produce ossigeno a partire da acqua e energia chimica a
partire da energia luminosa.
Fase oscura: Avviene nello stroma ed è anche
chiamata ciclo di Calvin. È fatto di un certo numero di reazioni e usa
anidride carbonica, ATP e NADPH. L'anidride carbonica si combina con un
composto a cinque atomi di carbonio detto ribulosio difosfato (RuDP). Questa
reazione è catalizzata da u enzima molto poco specifico detto rubisco
(ribulosio difosfato carbossilasi ossigenasi) poiché catalizza
indifferentemente la reazione tra il RuDP e l'anidride carbonica e la reazione
tra l'ossigeno e il RuDP. La prima tappa del ciclo consiste nella reazione tra
il ribulosio difosfato e l'anidride carbonica che produce un composto a tre
atomi di carbonio; seguono due tappe nelle quali viene utilizzato parte
dell'ATP prodotto dalla fase luminosa e tutte le molecole di NADPH per produrre
un composto a tre atomi di carbonio (gliceraldeide 3 fosfato). Nella terza
tappa due di qieste molecole escono dal ciclo consentendo la produzione di
glucosio. Le altre 10 che restano usando l'energia sottoforma di ATP servono a
ricostruire il ribulosio difosfato. Dal punto di vista energetico è un
processo un po' dispersivo.
Vari tipi di
fotosintesi: La grande
maggioranza delle piante eseguono il ciclo di Calvin sopra descritto e queste
piante sono dette C3. queste piante hanno però il problema
legato all'uso dell'enzima rubisco poiché quando la stagione è secca per
non disperdere acqua la pianta deve tenere gli stomi chiusi e il rubisco se
nella foglia c'è tanta anidride carbonica la catalizza altrimenti no e
quindi la reazione risulta sospesa. Se infatti il contenuto di anidride
carbonica è baso il rubisco catalizza la reazione con l'ossigeno
catalizzando la reazione di fotorespirazione e producendo glicolato. Ma
catalizzando questa reazione viene a mancare rubisco per catalizzare la
reazione con l'anidride carbonica. per evitare questo problema le piante C4
(canna da zucchero, mais) attuano un altro processo che avvia diversamente il
ciclo di Calvin. Non usano il rubisco ma un altro enzima che catalizza un'altra
reazione. L'enzima si chiama fosfoenol piruvato carbossilasi (PEP) e catalizza
la reazione fra anidride carbonica e fosfoenol piruvato, a differenza del
rubisco è altamente specifico per l'anidride carbonica e quindi
catalizza solo quella reazione. Alla fine di questa reazione si ottiene un
composto a 4 atomi di carbonio che si chiama acido ossalacetico o ossalacetato.
Questo composto viene fatto passare dentro le cellule della guaina vascolare
dove il quarto atomo di carbonio si stacca e si unisce di nuovo all'anidride
carbonica, rimane un composto a tre atomi di carbonio e il ciclo inizia
normalmente. Esistono anche piante CAM (metabolismo acido delle crassulacee),
le piante grasse. Si ha la stessa situazione delle piante C4 ma il
tutto avviene solo nelle cellule della guaina vascolare e di notte consentendo
agli stomi di restare chiusi per tutto il giorno. Le CAM sono quelle
maggiormente in grado di immagazzinare acqua anche perché hanno cellula - STRUTTURA DELLE CELLULE EUCARIOTE" class="text">delle cellule
in grado di assorbire molta acqua.