 |
|
| biologia |
|
GLI ACIDI NUCLEICI
Tutti gli organismi contengono acidi nucleici sotto forma di acido deossiribonucleico (DNA) e ribonucleico (RNA)
Il DNA è il depositario dell'informazione genetica che viene trascritta (cioè copiata) in molecole di RNA. L'RNA contiene il codice per sintetizzare specifiche proteine.
![]()
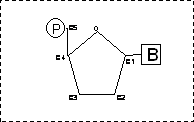 Una molecola acido nucleico è un polimero costituito da monomeri
detti nucleotidi. Ciascun nucleotide è costituito da tre molecole:
Una molecola acido nucleico è un polimero costituito da monomeri
detti nucleotidi. Ciascun nucleotide è costituito da tre molecole:
Uno zucchero pentoso è uno zucchero la cui molecola è costituita da 5 atomi di carbonio. La base azotata è legata al carbonio 1 dello zucchero pentoso, mentre l'acido fosforico (H3PO4) è legato al carbonio 5 che è esterno all'anello dello zucchero.
I nucleotidi si legano tra loro medianti legami fosfodiestere che collegano il C3 del pentoso di un nucleotide al C5 del nucleotide successivo. In questo modo l'acido fosforico impiega due dei suoi tre gruppi acidi nel legame fosfodiestere 3-5. il gruppo acido restante conferisce alla molecola di acido nucleico particolari caratteristiche:
Lo zucchero pentoso è il ribosio nell'RNA e il deossiribosio nel DNA. La differenza tra i due è che al deossiribosio manca l'ossigeno legato al C2.
Le basi azotate si chiamano così perché contengono molti atomi di azoto e possono essere di due tipi: purine o pirimidine
![]()
 BASI AZOTATE
BASI AZOTATE




![]()
I nucleotidi che costituiscono la molecola del DNA sono:
Quelli che costituiscono la molecola dell'RNA sono gli stessi, ad eccezione del timidin monofosfato che è sostituito dall'uridin monofosfato.
Un nucleoside è un nucleotide a cui manca il fosfato, cioè sono la combinazione di uno zucchero pentoso e di una base azotata. Per esempio, è un nucleoside l'adenosina:
Adenina → base azotata
Adenosina → nucleoside (adenina + zucchero pentoso)
Adenosin monofosfato → nucleotide (adenina + zucchero pentoso + fosfato)
L'adenosin trifosfato (ATP) è un particolare nucleotide con tre acidi fosforici, uniti tra loro con legami ad alto contenuto energetico. Infatti questa è una molecola fondamentale per la cellula perché rappresenta la principale forma di accumulo di energia.
La molecola di DNA è costituita da 2 catene polinucleotidiche le quali formano una doppia elica intorno allo stesso asse centrale. Esse sono antiparallele: i legami 3-5 fosfodiestere sono rivolti in direzione opposta. Questo vuol dire che se una catena inizia con il C3 libero e finisce con il C5 libero, l'altra catena è disposta in modo contrario.
Le due catene sono unite tra loro mediante legami a idrogeno che si instaurano tra le basi complementari. Le uniche coppie tra le quali è possibile il legame sono A-T e C-G.
Tra A e T si instaurano due legami a idrogeno mentre tra C e G se ne formano tre, quindi la coppia C-G è più stabile.
A=T C≡G
La sequenza assiale di basi lungo una catena può variare molto, ma la sequenza della catena corrispondente deve essere complementare alla prima.
Le due catene possono essere separate tra loro rompendo il legame a idrogeno tra le coppie di basi. Questo può essere fatto mediante riscaldamento o un pH alcalino (fusione o denaturazione). Dopo la denaturazione si può riottenere la conformazione a doppia elica lasciando raffreddare lentamente il DNA, in modo che le badi possano riappaiarsi (rinaturazione)
La struttura dell'RNA è simile a quella del DNA, tranne che per la presenza del ribosio invece del deossiribosio e dell'uracile invece della timina. Inoltre L'RNA è costituito da una catena singola.
Ma le molecole di RNA, avendo estese regioni complementari all'interno di una stessa catena, spesso si ripiegano e tra le basi della catena si instaurano legami a idrogeno, formando delle anse a forcina.
Ci sono tre tipi di RNA:


Nelle cellule degli eucarioti il DNA non è libero ma associato a piccole proteine dette istoni, in un complesso chiamato cromatina. Esistono cinque tipi differenti di istoni, tutti di natura basica. Per questo motivo possono instaurare uno stretto legame con il DNA, di natura acida.
I quattro istoni principali, H2A, H2B, H3 e H4 hanno ciascuno una composizione simile anche nelle specie più diverse, mentre l'istone H1 è differente da specie a specie.
La cromatina può presentare vari livelli di organizzazione:
Nel corso della divisione cellulare la cromatina si condensa a formare i cromosomi, i quali sono circa 40.000 volte più densi della fibra di 10 nm. Essi sono strutture bastoncellari che servono a poter distribuire equamente il DNA tra le cellule lie. I componenti dei cromosomi sono 4:
CROMATIDI alla metafase ciascun cromosoma risulta formato da due strutture simmetriche, i cromatidi, contenenti ognuna un'unica molecola di DNA. Essi sono esattamente uguali (cromatidi fratelli) e sono uniti fra loro a livello del centromero
CENTROMERO è la regione di attacco degli elementi del fuso mitotico sul cromosoma. È situato in un tratto più sottile del cromosoma detto costrizione primaria
TELOMERO estremità dei cromosomi contenenti l'inizio e la fine della molecola di DNA che costituisce il cromatidio
ORGANIZZATORE NUCLEOLARE si trovano in alcuni cromosomi che presentano costrizioni secondarie in cui ci sono regioni contenenti i geni che inducono la formazione del nucleolo
TEORIA UNINEMICA: ciascun cromatidio rappresenta un'unica molecola lineare di DNA con le proteine ad essa associate
I cromosomi vengono classificati in quattro tipi a seconda della forma determinata dalla posizione del centromero:
La cromatina si condensa nel corso della mitosi e si decondensa alla telofase. Ma ci sono alcune regioni dei cromosomi che non si decondensano e rimangono compatte anche durante l'interfase. Queste regioni vengono chiamate eterocromatine, mentre le altre eucromatina.
Nell'eterocramatina il DNA è molto denso e si presenta in forma di fibra di 20-30 nm cioè si tratta di DNA inattivo.
L'esempio più noto è quello della coppia di cromosomi X nelle femmine dei mammiferi: uno dei due cromosomi è attivo e rimane eucromatico mentre l'altro è inattivo e costituisce il corpo di Barr nell'interfase.
Un altro esempio è quello del gatto di Sna. Questo esemplare presenta un mantello a macchie nere e arancioni (a placche di tartaruga), ma solo le femmine. Questo perché la macchiatura un gene contenuto nel cromosoma X diventa eterocromatico e non attivo in alcuni gruppi di cellule e non in altri. In stadi precoci dell'embriogenesi, in ogni cellula della femmina di mammifero uno dei cromosomi X diventa inattivo a caso, di conseguenza nell'adulto si forma un mosaico in cui il 50% di cellule hanno un cromosoma X attivo di origine paterna e l'altro 50% un cromosoma X attivo di origine materna
IL CICLO CELLUARE
Una cellula in accrescimento presenta un ciclo cellulare che consiste essenzialmente in due periodi: l'interfase e la divisione. L'interfase presenta tre periodi che vengono chiamati fase G1,S e G2.
Duplicazione del DNA
La duplicazione del DNA avviene nella fase S (sintesi) del ciclo cellulare d'ogni cellula.
La replicazione del DNA è semi-conservativa: questo significa che ogni molecola lia contiene una catena parentale e una neo sintetizzata.
Il primo passo verso la replicazione semi-conservativa è la separazione delle due catene complementari nel punto in cui deve cominciare la replicazione; in modo che ciascuna catena sia libera di fungere da stampo per la polimerizzazione di una nuova catena complementare. La polimerizzazione è catalizzata dall'enzima DNApolimerasi; che seleziona i deossinucleotidi trifosfati (d-ATP, d-TTP, d-GTP, d-CTP) e li lega uno dopo l'altro con legame fosfodiestere 3'-5'. L'enzima lega il fosfato presente su un deossinucleotide trifosfato al gruppo OH legato al carbonio 3' del deossiribosio appartenente al nucleotide terminale della catena in crescita. Nella reazione i due gruppi fosfati terminali si staccano sotto forma di pirofosfato.
La DNApolimerasi catalizza solo l'allungamento unidirezionale di una catena, cioè può aggiungere nucleotidi solo all'estremità 3' ma non all'estremità 5'. La DNApolimerasi può catalizzare l'aggiunta di nucleotidi all'estremità 3' di una catena esistente ma non è in grado di iniziare una nuova catena. Per iniziarla è, infatti, necessario un innesco o primer cui aggiungere (all'estremità 3') i nucleotidi in sequenza. Nella cellula fa da innesco una breve catena di RNA. L'innesco è sintetizzato come segue:
prima avviene una dissociazione circoscritta delle due catene su un tratto interno alla molecola di DNA
un enzima detto primasi catalizza la formazione di una breve catena di RNA a partire da ribonucleotidi trifosfati (ATP, UTP, CTP, GTP) [la primasi a differenza della DNApolimerasi è capace di dare inizio ad una catena]
l'innesco di RNA resta appaiato allo stampo, poi arriva la DNApolimerasi che estende la catena per aggiunta di deossinucleotidi all'innesco
naturalmente si forma un innesco su ciascuna catena parentale e i due inneschi hanno polarità opposta e quindi le due catene lie crescono in direzioni opposte
man mano che le catene lie si estendono alla loro estremità 3' la doppia elica si apre a cerniera in entrambe le direzioni
il punto di rotolamento dell'elica si chiama forcella replicativi
man mano che la forcella avanza lascia dietro di se una zona a singolo filamento su ciascuna delle catene parentali. A questo punto si formano nuovi inneschi che sono poi prolungati per riempire a ritroso le parti non replicate delle catene parentali
gli inneschi sono poi rimossi quando i frammenti sintetizzati indietro (Okazaki) incontrano l'estremità 5' dell'innesco del tratto precedente. Quando avviene l'incontro la DNApolimerasi stacca uno dopo l'altro i nucleotidi dell'innesco e aggiunge simultaneamente deossinucleotidi
quando tutto l'innesco è stato rimosso le due estremità sono unite dalla DNAligasi
Nella replicazione del DNA bisogna considerare il problema che la separazione delle due catene richiede la despiralizzazione della doppia elica. La doppia elica fa un giro completo ogni 10 paia di basi. Quindi ogni 10 basi separate e svolte a livello della forcella replicativa, la doppia elica a valle deve fare un giro in direzione opposta.
In E.coli la forcella si apre alla V=60000 paia di basi al minuto, il che richiederebbe che la doppia elica a valle facesse 6000 giri al minuto (negli eucarioti la V è 10 volte inferiore). La rotazione di tutto il DNA a valle della forcella è bloccato dalle numerose proteine associate perciò in mancanza di rotazione, la torsione della doppia elica a valle ostacolerebbe la despiralizzazione e impedirebbe la replicazione. Il problema è stato risolto grazie all'enzima topoisomerasi che crea rotture transitorie in un singolo filamento a breve distanza dalla forcella replicativa. Queste rotture si creano e si riparano rapidamente ad opera della stessa topoisomerasi.
|
Privacy
|
© ePerTutti.com : tutti i diritti riservati
:::::
Condizioni Generali - Invia - Contatta
