|
|
| biologia |
|
|||||
|
I BATTERI
Sono microrganismi di natura cellulare poiché presentano alcune delle strutture tipiche delcellula - METABOLISMO, LA RESPIRAZIONE, RESPIRAZIONE AEROBICA DELLA SOSTANZA ORGANICA" class="text">la cellula, ma sono anche organismi molto semplici, poiché sono privi di alcune delle strutture tipiche delle cellule eucariotiche.
Hanno dimensioni dell'ordine di grandezza del μm, per cui sono visibili anche al microscopio ottico.
Possono assumere varie forme, in particolare possono essere cocchi (a forma sferica), bacilli (a forma di bastoncino), spirilli (a forma elicoidale). I batteri possono poi aggregarsi in varie strutture che prendono nomi diversi a seconda della loro morfologia: diplococchi (associazione di due batteri), streptococchi (i batteri si associano per formare una catenella), stafilococchi (i batteri si associano costituendo una particolare struttura a grappolo).
I batteri presentano poi alcune strutture essenziali per la loro esistenza ed altre strutture accessorie.
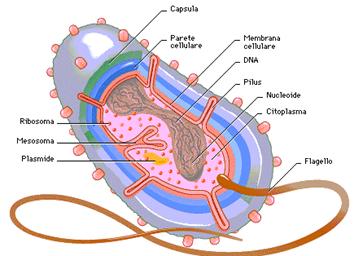
Fra le strutture essenziali sono da ricordare:
Il DNA, raccolto in unico cromosoma a doppia elica circolare, che nella cellula si ritrova in forma superavvolta. Alcuni tratti possono essere legati alla membrana plasmatica; ciò ha senso dal momento che a questo livello si localizzano tutti gli enzimi necessari per il metabolismo cellulare, compresi quelli per la replicazione e la trascrizione del DNA. Si hanno poi sequenze geniche separate dal cromosoma, chiamate plasmidi; tali strutture sono importanti dal momento che sono proprio esse le parti di genoma batterico ad essere trasferite durante la riproduzione sessuata ed è proprio la variabilità conferita da queste strutture a determinare, in alcuni casi, la formazione di ceppi resistenti ai farmaci o ai disinfettanti.
Si hanno poi i ribosomi, diversi da quelli delle cellule eucariotiche per via di sostanziali differenze nella composizione, sia per quanto riguarda l'RNA ribosomiale, sia per quanto riguarda la componente proteica. Il loro coefficiente di sedimentazione è di 70s, con la subunità minore di 30s e la maggiore di 50s. Si trovano liberi nel citoplasma.
La membrana plasmatica batterica è molto simile a quella delle cellule eucariotiche dal momento che anch'essa è costituita da fosfolipidi e proteine con funzioni di trasporto ed enzimatiche. L'unica differenza sta nel fatto che nella membrana plasmatica batterica mancano gli steroli.
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() La parete è l'ultima delle
strutture essenziali. È rigida e ciò le permette di avere due
importanti funzioni: la prima è quella di impedire la lisi della cellula
a causa di un eccessivo assorbimento d'acqua, la seconda è quella di
proteggerla da agenti esterni pericoloso i tossici. La rigidità è
conferita da una particolare sostanza chiamata peptidoglicano, la quale
è un polimero formato da monomeri fra di loro simili. Ciascun monomero
è costituito da due ammino-zuccheri chiamati NAM e NAG; al NAM è
legata una corta catena di 5 aminoacidi in cui il primo è sempre una
L-Ala, mentre il quarto ed il quinto sono D-Ala. NAM e NAG sono legati fra loro
a costituire il monomero per mezzo di un legame di tipo β1-4. Ciascun
monomero è poi legato al successivo per mezzo di un legame β1-6.
La parete è l'ultima delle
strutture essenziali. È rigida e ciò le permette di avere due
importanti funzioni: la prima è quella di impedire la lisi della cellula
a causa di un eccessivo assorbimento d'acqua, la seconda è quella di
proteggerla da agenti esterni pericoloso i tossici. La rigidità è
conferita da una particolare sostanza chiamata peptidoglicano, la quale
è un polimero formato da monomeri fra di loro simili. Ciascun monomero
è costituito da due ammino-zuccheri chiamati NAM e NAG; al NAM è
legata una corta catena di 5 aminoacidi in cui il primo è sempre una
L-Ala, mentre il quarto ed il quinto sono D-Ala. NAM e NAG sono legati fra loro
a costituire il monomero per mezzo di un legame di tipo β1-4. Ciascun
monomero è poi legato al successivo per mezzo di un legame β1-6.
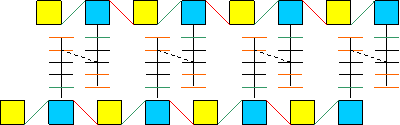
La parete è poi costituita da più strati di peptidoglicano; il legame fra i vari strati è consentito da legami fra gli aminoacidi delle catene aminoacidiche legate a NAM: il terzo aminoacido della catena legata ai NAM del polimero superiore è legato covalentemente al quarto aminoacido della catena legata ai NAM del polimero inferiore. La formazione di tale legame è catalizzata dall'enzima transpeptidasi, mentre l'energia necessaria affinché la reazione avvenga è fornita dall'idrolisi del legame peptidico fra il quarto ed il quinto aminoacido di ciascuna catena.
La natura della parete cellulare è però diversificata a seconda che si parli di batteri Gram+ e batteri Gram-.
Nei batteri Gram+ il legame fra il terzo aminoacido di una catena ed il quarto dell'altra è intervallato da cinque Gly, per cui la parete dei Gram+ è più spessa di quella dei Gram-. Fra le maglie della loro parete, i Gram+ presentano degli acidi teoicoici, la cui funzione è quella di legare alla superficie dei batteri degli ioni positivi, indispensabili per il loro metabolismo.
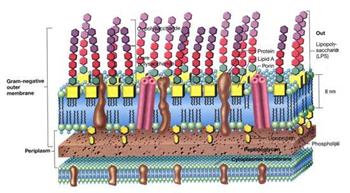 Nei batteri Gram-, invece, il legame fra le catene aminoacidiche
è diretto e e una struttura assente nei Gram+, la membrana
esterna.
Nei batteri Gram-, invece, il legame fra le catene aminoacidiche
è diretto e e una struttura assente nei Gram+, la membrana
esterna.
Questa struttura è un doppio foglietto la cui parte interna è di natura lipidica, mentre la parte esterna è di natura lipopolisaccarididica (LPS).
Il foglietto esterno è a sua volta formato da tre strati: internamente si ha il lipideA, l'endotossina di questi batteri, intermediamente si ha lo strato C, di natura saccaridica e uguale per tutti i Gram-, mentre esternamente si ha l'antigene O, di natura saccarifica.
In questa membrana sono poi immerse delle proteine chiamata porine, che permettono il passaggio di piccole molecole idrofile, che altrimenti non riuscirebbero a passare questa barriera.
Fra le strutture non essenziali sono da ricordare:
La capsula, una struttura di natura polisaccaridica presente in batteri Gram+ e Gram- che funge da adesina (permette al batterio di aderire alle superfici o ai tessuti) o possiede attività antifagocitaria; inoltre permette ai batteri di diventare immuni agli antibiotici.
I flagelli sono formati da una proteina chiamata flagellina e sono organi che permettono la motilità del batterio ruotando in senso orario od antiorario al fine di avvicinarlo a sostanze nutritizie (chemiotassi positiva) o allontanalo da sostanze pericolose (chemiotassi negativa).
I pili sono appendici più corte dei flagelli che hanno funzione di ancoraggio ai tessuti o ad altri batteri, particolari pili sono i pili sessuali, che permettono lo scambio di materiale genetico fra batteri.
Condizioni di crescita dei batteri
I batteri possono essere o autotrofi (traggono il loro nutrimento per mezzo della fotosintesi) o eterotrofi (traggono il loro nutrimento assorbendolo dall'esterno).
Le condizioni di crescita dei batteri possono essere molto variabili, ma a seconda del tipo di batterio è comunque necessario regolare alcuni parametri specifici:
acqua: fondamentale per tutti gli organismi;
pH: compreso fra 6 e 8;
temperatura: compresa fra i 30 e 45°C per i mesofili; alta per i termofili; bassa per i psicrofili.
pressione osmotica: la concentrazione salina del mezzo esterno ottimale dovrebbe essere circa del 0,85%, ma alcuni batteri possono sopravvivere anche a concentrazioni del 10%.
ossigeno: alcuni batteri sono anaerobi obbligati, cioè sono costretti a vivere in assenza di ossigeno; altri sono aerobi facoltativi, cioè possono vivere in ambienti con o senza ossigeno; altri ancora sono aerobi obbligati, ovvero possono vivere solo in presenza di ossigeno.
Moltiplicazione batterica e formazione di spore
I batteri si moltiplicano per scissione binaria: il cromosoma batterico viene addossato alla membrana plasmatica, dove gli enzimi specifici lo replicano; a questo punto le due molecole di DNA passano ai poli della cellula e si formano internamente dei setti membranosi; i setti poi si uniscono e viene formata su di essi la parete cellulare.
In laboratorio è molto spesso necessario effettuare la conta dei batteri, che può essere fatta sia su batteri vivi, sia su batteri morti.
Per quanto riguarda la conta dei batteri morti, essa può essere fatta seguendo diverse procedure:
La conta diretta al microscopio, servendosi di particolari contenitori chiamati contenitori portacellule,entro cui vengono messi i batteri, oppure la conta per densità ottica, ovvero facendo passare fasci di luce attraverso la coltura e contando quelli che non vengono deviati (gli altri vengono deviati dalle cellule batteriche).
I batteri vivi vengono invece contati per mezzo di diluizioni successive della coltura, considerando che ogni colonia è nata da un singolo batterio.
La conta dei batteri è utile per conoscere il numero di batteri presenti in una colonia in funzione del tempo. Dai dati raccolti è poi possibile ottenere una curva, chiamata curva di crescita.
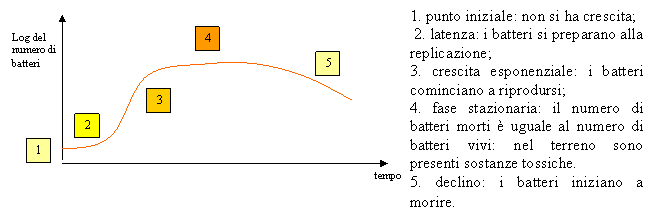
Nel momento in cui le condizioni dell'ambiente diventano avverse per la vita del batterio, vengono sviluppate delle forme di resistenza, chiamata spore.
Le spore hanno origine dalla cellula vegetativa e sono metabolicamente inerti, poiché quasi completamente disidratate: l'acqua viene invece sostituita dall'acido diticolinico associato a ioni calcio.
Esosporio
 È formata da diversi involucri: all'interno si ha il protoplasto, contenente gli acidi
nucleici, le proteine e quanto rimane del citoplasma. Il protoplasto è
poi circondato da residui della membrana plasmatica e della parete cellulare,
mentre all'esterno si hanno la corteccia,
formata da peptidoglicano modificato, e due tuniche chiamate tunica interna e tunica esterna, composte da proteine solforate (maggiormente
stabili); più esternamente ancora, si ha l'esosporio, di natura fosfolipoproteica.
È formata da diversi involucri: all'interno si ha il protoplasto, contenente gli acidi
nucleici, le proteine e quanto rimane del citoplasma. Il protoplasto è
poi circondato da residui della membrana plasmatica e della parete cellulare,
mentre all'esterno si hanno la corteccia,
formata da peptidoglicano modificato, e due tuniche chiamate tunica interna e tunica esterna, composte da proteine solforate (maggiormente
stabili); più esternamente ancora, si ha l'esosporio, di natura fosfolipoproteica.
Nel genoma della cellula batterica si hanno particolari geni che regolano lo stato di spora e che vengono attivati nel momento in cui le condizioni ambientali diventano difficili per il batterio: l'attivazione avviene per mezzo del cambiamento conformazionale indotto dai cambiamenti ambientali sulla DNApolimerasi, che così modificata riconosce unicamente i geni per lo stato di spora.
Il passaggio da cellula vegetativa a spora (sporulazione) avviene secondo alcune fasi:
il genoma batterico viene copiato interamente;
una delle due molecole di DNA formatesi si sposta ad un polo della cellula;
attorno al DNA che si è spostato al polo si formano gli involucri sporali;
la cellula contenente la spora (sporangio) va in lisi liberando la spora nell'ambiente.
Il passaggio da spora a cellula vegetativa (germinazione) avviene nel momento in cui le condizioni tornano ad essere favorevoli:
si rompono i mantelli sporali permettendo l'ingresso dei metabolici;
cominciano ad essere degradati i mantelli e viene eliminato l'acidi diticolinico, mentre si ha l'idratazione della cellula;
riprende il metabolismo respiratorio della cellula;
la nuova cellula riprende a moltiplicarsi.
Genetica dei batteri
Il genoma batterico presenta alcune caratteristiche:
Mancano gli introni, per cui tutte le sequenze vengono trascritte e codificano per proteine.
La replicazione è semiconservativa ed avviene con i medesimi meccanismi attuati nelle cellule eucariotiche;
Il genoma batterico è raccolto in un unico cromosoma costituito da DNA a doppia elica circolare. La replicazione di tale genoma ha come punto di origine O, da cui poi prende origine la "bolla d replicazione".
Si hanno tre topoisomerasi, che svolgono funzioni differenti: la prima provoca la despiralizzazione del cromosoma, la seconda la spiralizzazione, la terza provoca la decatenazione.
I filamenti di mRNA spesso codificano per più proteine e mancano di CAP e sequenza di poliA. Inoltre giungono immediatamente ai ribosomi, dal momento che il DNA batterico si trova immerso nel citoplasma.
La maggior parte delle proteine batteriche comincia con l'amminoacido metionina, convertito normalmente in formil-metionina, proteina riconosciuta da macrofagi e linfociti.
Il controllo della traduzione è dato da particolari geni chiamati operon, che codifica per enzimi costitutivi, sempre presenti nella cellula, ed enzimi inducibili, prodotti solamente quando il substrato è presente.
Il DNA batterico viene replicato allo stesso modo del DNA eucariotico, discorso diverso vale per i plasmidi.
In corrispondenza dell'origine O una delle due catene viene tagliata e la polimerasi3 comincia ad aggiungere nucleotidi in direzione 5'→ 3'.
L'aggiunta di nucleotidi fa ruotare la molecola di DNA in senso antiorario, mentre l'estremità 5' si linearizza, formando una coda libera.
Con il procedere della sintesi, si forma una molecola lia di DNA a doppia elica circolare e una molecola lia di DNA a doppia elica lineare.
Infine anche la molecola lia lineare si circolarizza.
Questo meccanismo è stato ideato per spingere il plasmide replicato in un batterio lio o in un batterio ricevente.
I plasmidi sono molecole di DNA esterne al cromosoma batterico, che non codificano per proteine essenziali per la sopravvivenza del batterio, ma per tossine, adesine, enzimi responsabili della resistenza ai farmaci e batteriocine (proteine che uccidono i batteri di altre specie).
All'interno vi si trovano sequenze chiamate TRA, che permettono il trasferimento dei plasmidi da una cellula ad un'altra, sequenze responsabili della replicazione autonoma del plasmide e sequenze di inserzione, per inserirsi nel cromosoma batterico.
I batteri hanno la possibilità di scambiarsi o trasferire materiale genetico da una cellula all'altra o all'interno della stessa cellula da un sito all'altro. Questo trasferimento avviene per mezzo di alcuni meccanismi specifici:
Trasformazione: in questo caso, il genoma di un batterio morto, che rimane ancora attivo, viene in contatto con quello di un batterio vivo e lo modifica, creando così un nuovo ceppo.
Per prima cosa, si ha l'attacco del genoma al batterio ricevente nel momento in cui quest'ultimo è competente, ovvero nel momento in cui il suo livello di sintesi proteica è massimo e quello di DNA è minimo. Il genoma, quindi, penetra nella cellula e va in eclisse, cioè diventa non-trasformante dal momento che uno dei due filamenti della doppia elica è stato digerito. Dopo la riparazione di questo filamento, si ha la ricombinazione e la sa del nuovo fenotipo.
Durante la competenza, la cellula batterica sintetizza anche particolari proteine chiamate fattori di competenza, che si legano a recettori di membrana specifici, attivando la sintesi delle autolisine, proteine che producono dei fori nella parete cellulare consentendo così l'aggancio di altre proteine che legano il DNA ricombinante e ne digeriscono un filamento.
 Coniugazione: nei batteri Gram-, cellule
donatrici, chiamate F+ , presentano un plasmide, nel quale si
trovano geni per la sintesi del pilo sessuale, geni per il trasferimento del
DNA e geni per la replicazione autonoma del plasmide. Le cellule riceventi F-
sono invece prive di questo plasmide.
Coniugazione: nei batteri Gram-, cellule
donatrici, chiamate F+ , presentano un plasmide, nel quale si
trovano geni per la sintesi del pilo sessuale, geni per il trasferimento del
DNA e geni per la replicazione autonoma del plasmide. Le cellule riceventi F-
sono invece prive di questo plasmide.
Nel momento in cui donatore e ricevente entrano in contatto per mezzo del pilo sessuale, il donatore replica il plasmide e la molecole lia lineare viene inserita nel canale di coniugazione per raggiungere la cellula ricevente. Al termine del processo si avranno quindi solamente cellule F+.
Talvolta, il plasmide si integra all'interno del cromosoma batterico, per cui, assieme ad esso, può essere trasferito l'intero cromosoma. In questi casi, però, non è detto che la cellula ricevente diventi F+, dal momento che questo trasferimento è molto lungo ed il pilo in questo lasso di tempo può rompersi, impedendo quindi l'intero trasferimento del cromosoma ed eventualmente del plasmide F in esso integrato.
Nei batteri Gram+, invece, il contatto fra cellule non avviene per mezzo del pilo sessuale, ma per mezzo di proteine specifiche secrete dal donatore.
Nei plasmidi, oltre al fattore F, possono ire altri fattori come i fattori R, che conferiscono resistenza ai farmaci dal momento che sintetizzano enzimi che bloccano i principi attivi, fattori col, che permettono la sintesi di colicine, sostanze che uccidono i batteri competitori, fattori ent, che producono enterotossine.
 Trasduzione: è
un meccanismo mediato da un virus che infetta i batteri (batteriofago, virus a DNA con
testa icosaedrica, collare, coda a struttura spiralizzata, corona ad uncini e
fibre). In particolare, per questo tipo di trasferimento, sono utili i fagi
temperati, ovvero quelli il cui genoma si integra in quello batterico e solo in
seguito ad opportuni stimoli cominciano a moltiplicarsi, dando così un
ciclo lisogeno.
Trasduzione: è
un meccanismo mediato da un virus che infetta i batteri (batteriofago, virus a DNA con
testa icosaedrica, collare, coda a struttura spiralizzata, corona ad uncini e
fibre). In particolare, per questo tipo di trasferimento, sono utili i fagi
temperati, ovvero quelli il cui genoma si integra in quello batterico e solo in
seguito ad opportuni stimoli cominciano a moltiplicarsi, dando così un
ciclo lisogeno.
Si distinguono quindi una trasduzione generalizzata e una traduzione specifica.
Nella traduzione generalizzata, durante la fase d'assemblaggio, la testa del fago acquisice un frammento, di dimensioni opportune, di DNA batterico. Con la lisi della cellula, quindi, questi fagi trasducenti potranno attaccare altre cellule provocando l'inserimento di nuovi geni. Si parla di traduzione generalizzata dal momento che l'inserimento del frammento di DNA è del tutto casuale.
Nella traduzione specifica, invece, il DNA fagico per riattivarsi ha bisogno di essere exciso dal DNA batterico; questo "taglio", però, può non essere preciso e possono essere inglobati anche geni batterici, portando quindi alla formazione di un DNA virale ibrido. A questo punto, comincia il ciclo litico; se il DNA virale è sufficiente per formare virioni maturi, verrà integrata anche la parte di DNA batterico che potrà integrarsi nel batterio-ospite che sarà infettato dal virione formatosi. In questo caso si parla di specificità dal momento che vengono inglobati solamente i geni batterici adiacenti al genoma virale.
4. Trasposizione: in un batterio esistono sequenze poste sul cromosoma o sul plasmide chiamate elementi trasponibili. Questi elementi hanno la capacità di spostarsi da una zona all'altra della molecola di DNA o di passare da una molecola di DNA ad un'altra.
Affinchè si abbia la trasposizione, le sequenze trasponibili devono essere copiate; in questo modo si avranno due sequenze identiche in due punti diversi della molecola.
Gli elementi trasponibili possono essere di tre tipi: sequenze di inserzione (tratti piccoli formati da brevi sequenze ripetute ed invertite che codificano per l'enzima deputato alla trasposizione e alla duplicazione della sequenza); trasposoni (sequenze più complesse alle cui estremità si hanno le sequenze di inserzione); elementi invertibili (sono simili ai trasposoni, ma in questi casi è presente l'enzima invertasi che ha il compito di invertire l'orientamento della sequenza).
Patogenesi batterica
Generalmente i atteri sono parassiti, che possono vivere da simbionti, da commensali o da patogeni all'interno dell'ospite.
Per tale motivo, è necessario distinguere tra infezione e malattia, dal momento che spesso, la presenza e la riproduzione dei batteri, ovvero l'infezione, è del tutto asintomatica, mentre in altri casi porta alla sa di sintomi clinici, ovvero alla malattia.
La patogencità di un batterio è provocata da alcuni fattori come la sua capacità di moltiplicarsi nell'ospite e la capacità di produrre particolari sostanze tossiche responsabili della sa dei sintomi.
Generalmente i primi siti di infezione sono la mucosa enterica o le prime vie respiratorie; a questo livello i batteri aderiscono per mezzo di sostanze di natura proteica chiamate adesine e cominciano a riprodursi costituendo un biofilm, utile perché permette alla colonia di essere meno attaccabile dal sistema immunitario.
La moltiplicazione può essere favorita da alcuni fattori come l'attività antifagocitaria, conferita da alcune strutture batteriche o da alcune sostanze che allontanano o uccidono le cellule del nostro sistema immune, permettono di ricoprire il batterio di fibrina, o degradano H2O2, ovvero la sostanza impiegata dai macrofagi per digerire gli elementi estranei.
Durante la colonizzazione, i batteri cominciano a secernere sostanze tossiche chiamate agressine (che favoriscono la penetrazione dei batteri nei tessuti più profondi) e tossine.
Fra le agressine, sono da ricordare la collagenasi, che degrada le fibre collagene presenti nel connettivo e nei muscoli, la ialorunasi, che degrada l'acido ialuronico presente nel connettivo, la lipasi, che attacca i lipidi, la chinasi, che degrada i coaguli di fibrina che si formano durante i processi di infiammazione, la proteasi, che degrada le proteine, e la nucleasi, che degrada gli acidi nucleici.
Le tossine devono essere invece suddivise in endotossine ed esotossine.
Le endotossine sono di natura lipidica, derivano infatti dal lipideA, e sono presenti unicamente nei batteri Gram-. La loro azione è generalizzata, così come è generalizzato il quadro clinico che provocano.
Le esotossine sono invece di natura proteica e sono presenti sa in batteri Gram+, sia in batteri Gram-. La loro azione, di natura enzimatica, è estremamente specifica ed anche la sintomatologia ad esse connesse è particolarmente peculiare.
Esotossine: ogni esotossina ha un suo bersaglio specifico ed un preciso meccanismo d'azione. Generalmente le tossine sono proteine dimeriche in cui si riconoscono due subunità distinte, una subunità A di natura enzimatiche che interferisce con il metabolismo cellulare provocando gli effetti della tossina ed una subunità B che funge da antirecettore e riconosce la cellula-bersaglio. Le due subunità sono fra loro legate da ponti disolfuro e, una volta entrate nella cellula, vengono ridotte in modo da dividersi nelle due parti funzionali.
Tossine con azione diretta sulla membrana: due esempi sono la tossina epidermolitica e le citolisine. Entrambe vengono prodotte da Stafilococcus aurus. La prima agisce a livello dello strato granuloso dell'epidermide provocando la rottura delle proteine che compongono la matrice intracellulare, il che si traduce nella "sindrome della cute ustionata", mentre la seconda agisce sulla membrana plasmatica provocando la formazione di canali e quindi una massiccia entrata di acqua, che porta a lisi osmotica della cellula.
Tossine che agiscono in seguito alla penetrazione nella cellula: esiste un gruppo molto ampio di tossine dette ADP-ribosillanti, che alterano alcuni meccanismi di segnalazione intercellulare poiché agiscono sulle proteine G: agiscono infatti sulle proteine Gs mantenendole sempre attive e portando ad una produzione massiccia di cAMP, ciò avviene perché tali tossine agiscono sul NAD staccando da esso l'adenosil-difosfato-ribosio e legandolo alle proteine G. Esempi di queste tossine sono la tossina del colera, la tossina della pertosse e la tossina difterica.
La tossina del colera è formata da 5 subunità B e una subunità A, divisa in A1 e A2. Le subunità B si legano a gangliosidi presenti sulla membrana degli enterociti dell'intestino tenue. Questo legame favorisce l'entrata della subunità A nella cellula, che al suo interno si divide nelle subunità A1 e A2. la parte A1 provoca un aumento di cAMP con conseguente perdita di acqua ed elettroliti da parte della cellula.
La tossina della pertosse viene liberata a livello della trachea e dei bronchi e poi, per mezzo della circolazione sanguigna, raggiunge varie cellule, fra cui quelle del pancreas e della tiroide, provocando la liberazione dell'insulina e di altre sostanze che portano a ipotensione e vasodilatazione. Ciò è dovuto al fatto che questa tossina interagisce con la proteina Gi che non riceve più stimoli inibitori e non modula più la produzione di cAMP, che diventa quindi eccessivamente concentrato.
La tossina della difterite è codificata dal gene TOX, presente nei profagi. L'infezione difterica si localizza a livello della prime vie respiratorie, mentre la tossina, per mezzo della circolazione sanguigna, diffonde nell'intero organismo. È divisa in due subunità, unite da ponti disolfuro, che vengono separate dopo l'endocitosi. La subunità A va ad interagizire con il fattore di inizio EF2, che innesca l'inizio della sintesi proteica. In questo modo la sintesi viene bloccata e la cellula muore.
Tossine neurotrope: sono tossine che bloccano la trasmissione degli impulsi nervosi:
Tossina del tetano: il tetano è un batterio che vive nel terreno sottoforma di spora e viene riattivato nel momento in cui viene a contatto con le ferite. A questo punto il batterio si replica e comincia a produrre la tossina, che si lega a siti neuronali specifici, bloccando l'impulso inibitore da stiramento (quando noi contraiamo un muscolo, contemporaneamente altri muscoli vengono stirati e mandano impulsi al cervello; per avere movimenti corretti è però necessario inibire queste contrazioni sinergiche). Uno dei sintomi classici dell'infezione da tetano è infatti la paralisi spastica, dovuta al fatto che i muscoli rimangono sempre contratti. A livello molecolare, l'azione è dovuta all'interazione fra subunità A della tossina ed proteina sinaptobrevina, localizzata sulle vescicole che trasportano i neurotrasmettitori regolandone il trasporto; quest'interazione provoca il blocco del passaggio dell'impulso nervoso da neurone a neurone.
Tossina del botulino: questa tossina agisce a livello delle sinapsi neuro-muscolari, bloccando il rilascio dell'acetilcolina, responsabile della trasmissione dell'impulso nervoso da neurone a muscolo. Il botulino è un batterio che può contaminare i cibi mal sterilizzati e infetta l'intestino, mentre la tossina va in circolo, colpendo il sistema nervoso.
Tossine che agiscono come superantigeni: normalmente gli antigeni, sostanze estranee, vengono fagocitate da cellule specifiche come i macrofagi e quindi presentate ai linfociti T, che riconosceranno l'antigene e daranno la risposta immune specifica. La risposta immune deve però essere bilanciata, in modo da essere efficace, ma non pericolosa per l'organismo. Le tossine che agiscono da superantigene sono tossine capaci di stimolare il sistema immune senza essere presentate dai macrofagi, ma si legano in maniera aspecifica a tutte le cellule del sistema immunitario provocando una massiccia produzione di citochine, necessarie per la comunicazione fra le cellule del sistema immune; questa spropositata risposta immune porta quindi a vari sintomi, come nausea, vomito, febbre e altri sintomi.
Tossine ciliostatiche: bloccano i movimenti ciliari e provocano la necrosi degli epiteli ciliati, come quelli della mucosa intestinale o delle vie respiratorie. Questa condizione permette quindi una maggiore possibilità di sviluppo dell'infezione batterica.
Eendotossine: hanno struttura analoga per tutti i batteri Gram-, hanno come unico precursore chimico il lipideA e presentano quindi tutte lo stesso meccanismo d'azione.
Normalmente le endotossine sono presenti nel nostro organismo e, a piccole dosi, sono benefiche poiché stimolano il sistema immune; la patogenicità deriva da una massiccia presenza di queste molecole, che provocano febbre (le tossine raggiungono l'ipotalamo, dove stimolano la produzione di prostaglandine, che hanno il compito di regolare la temperatura corporea; un eccesso di prostaglandine porta ad un innalzamento della temperatura corporea), formazione di trombi e vasodilatazione.
Siccome sono frequenti contaminanti di vari prodotti, è stato ideato un test per individuarle, il limulus test, che si serve dell'endolinfa di un crostaceo, che ha la proprietà di gelificare in presenza di endotossine.
Farmaci antibatterici
I farmaci antibatterici si dividono in due grandi classi: i chemioterapici e gli antibiotici. Questi farmaci, diversamente dai disinfettanti, vanno ad agire su momenti specifici del metabolismo del batterio. Possono poi dividersi in due categorie: i batteriostatici, che bloccano la replicazione dei batteri senza ucciderli, rendendoli così più vulnerabili all'attacco del sistema immunitario, e i battericidi, che uccidono le cellule batteriche.
Chemioterapici: i chemioterapici sono farmaci artificiali.
Sulfamidici: agiscono come antimetaboliti, dal momento che sono competitori con il substrato dell'enzima diidropteroato-sintetasi. Questo enzima fa parte del metabolismo dei derivati dell'acido folico, che fungono da donatori di unità carboniose nella sintesi di acidi nucleici ed aminoacidi. Questi farmaci sono poi selettivi, dal momento che le cellule umane assumono l'acido folico dalla dieta e non esistono vie metaboliche al loro interno. Questi farmaci non sono utili nelle infezioni da enterococco, poiché questo batterio assorbe l'acido folico dall'ambiente circostante, e nel caso di infezioni con necrosi cellulare o tissutale, dal momento che in questi ambienti la sintesi dell'acido folico è inutile per i batteri dal momento che l'ambiente è comunque ricco di aminoacidi ed acidi nucleici.
Isoniazide: è un analogo della nicotin-ammide ed inibisce la sintesi del NAD; inoltre è un analogo della vitamina B6 per cui può inibire l'azione di tutti quegli enzimi che hanno come cofattore questa vitamina.
Chinoloni: derivano dall'acido nalidixico e inattivano la subunità A della topoisomerasi II, responsabile del taglio della doppia elica di DNA in siti specifici. Esiste poi un antibiotico, la Novobiocina, che agisce sulla subunità B, per cui chinoloni e Novobiocina sono spesso impiegati sinergicamente.
Antibiotici: sono farmaci naturali, prodotti da batteri e miceti coltivati opportunamente. Possono essere distinti in classi a seconda del loro meccanismo d'azione.
Antibiotici che agiscono sulla sintesi della parete cellulare: la sintesi della parete cellulare inizia a livello citoplasmatico, dove il NAG viene condensato con il fosfoenolpiruvato e successivamente ridotto ad acido lattico e NAM, al NAM vengono poi legati i cinque aminoacidi per mezzo dell'enzima transpeptidasi. A questo livello della sintesi della parete agisce la cicloserina, che impedisce la trasformazione della L-Ala in D-Ala e di conseguenza la formazione del dimero di D-Ala.
A questo punto le molecole di NAM e NAG sintetizzate vengono legate al bactoprenolo, un trasportatore di natura lipidica, e portate a livello della parete cellulare: il bactoprenolo, cui è legato un gruppo fosfato, lega in NAM-pentapeptide; al NAM viene poi legato il NAG, per mezzo dell'enzima transglicosilasi, e si forma quindi il monomero. Il bactoprenolo in questo momento del ciclo ha legato un secondo gruppo fosfato. In prossimità della parete il monomero viene rilasciato e, grazie ad opportuni enzimi, viene legato al polimero di peptidoglicano in via di formazione. Questi enzimi, chiamati PBP, sono il target d'azione della penicillina, che agisce come analogo strutturale delle due D-Ala e rende inattivo il sito attivo dell'enzima transpeptidasi. Si hanno poi la vancomicina, che impedisce che il monomero si stacchi dal bactoprenolo, e la bacitraccina, che impedisce che il bactoprenolo, fosforillato alla fine del ciclo, possa essere defosforillato e ritorni attivo.
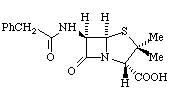 Nella penicillina, il nucleo fondamentale è l'acido 6-amminopenicillanico, a sua volta costituito
da un anello βlattamico e un anello tiazolidinico. All'anello
βlattamico è legato un gruppo amminico, importante perché a questo
livello possono essere legati diversi radicali che conferiscono così
alla molecola proprietà differenti. L'anello βlattamico può
poi essere degradato da enzimi specifici chiamati βlattamasi; questi
enzimi, che non sono presenti in tutti i batteri, costituiscono un fattore di
resistenza alla penicillina.
Nella penicillina, il nucleo fondamentale è l'acido 6-amminopenicillanico, a sua volta costituito
da un anello βlattamico e un anello tiazolidinico. All'anello
βlattamico è legato un gruppo amminico, importante perché a questo
livello possono essere legati diversi radicali che conferiscono così
alla molecola proprietà differenti. L'anello βlattamico può
poi essere degradato da enzimi specifici chiamati βlattamasi; questi
enzimi, che non sono presenti in tutti i batteri, costituiscono un fattore di
resistenza alla penicillina.
La penicillina è attiva solamente sui batteri Gram+ per cui tutte le penicilline sintetiche vertono ad essere ad ampio spettro (devono agire anche sui batteri Gram-) e resistenti alle βlattamasi. Di solito, però, i farmaci sintetici tendono ad avere solo una di queste due caratteristiche; ad esempio la ampicillina è ad ampio spettro, ma non è resistente alle βlattamasi; la oxacillina non è ad ampio spettro, ma è resistente alle βlattamasi.
Le cefalosporine hanno un meccanismo d'azione simile a quello della penicillina (come la penicillina, possiedono l'anello βlattamico), ma hanno il vantaggio di essere ad ampio spettro e di resistere alle βlattamasi; nonostante ciò sono meno consigliabili delle penicilline.
Antibiotici che agiscono sulle membrane plasmatiche: i principali sono le pimixine, attive sui Gram- perché s legano allo strato di LPS, che compone la loro membrana esterna.
Antibiotici che agiscono sulla sintesi degli acidi nucleici: due esempi sono la novobiocina e la rifomicina; il primo farmaco si lega alla girasi batterica e per questo agisce in sinergia con i chinoloni; il secodo è un inibitore della sintesi dell'RNA ed è ad ampio spettro e selettivo, dal momento che la RNApolimerasi batterica è diversa da quella umana.
Antibiotici che agiscono sulla sintesi proteica: gli antibiotici che agiscono sulla subunità 30s (tetracicline e aminoglicosidi, come la gentamicina, la neomicina e la streptomicina) agiscono legandosi alla subunità, sottraendola al pool ribosomiale ed impedendo la formazione del ribosomi completi. Agiscono in sinergismo con gli anelli βlattamici. Gli antibiotici che agiscono sulla subunità 50s, come i macrolidi o il cloramfenicolo, bloccano la formazione del ribosomi completo e, dal momento che hanno azione analoga, non possono essere utilizzati in sinergia fra loro.
La resistenza ai farmaci è una caratteristica di alcuni batteri, che viene in genere acquisita per mezzo di mutazioni genetiche; le mutazioni possono provocare cambiamenti di vario tipo, ad esempio possono essere sviluppati enzimi che degradano il principio attivo, oppure vengono modificati i bersagli del farmaco, o ancora, può diminuire la permeabilità al farmaco, infine, possono essere sviluppate vie metaboliche alternative a quelle che il farmaco va a bloccare.
|
Privacy
|
© ePerTutti.com : tutti i diritti riservati
:::::
Condizioni Generali - Invia - Contatta