 |
|
| biologia |
|
|
|||||
|
Il Controllo Dell'Espressione Genica
Quando un gene è attivato, viene trascritto in RNA e il suo messaggio è tradotto in specifiche molecole proteiche. Il processo complessivo con cui l'informazione genetica passa dai geni alle proteine è detto "espressione genetica". Ad esempio l'E.Coli può trovarsi nel nostro intestino in ambienti diversi: ambiente ricco di grassi e glucosio, se si è mangiato pane e burro, o di lattosio, se si è bevuto un bicchiere di latte scremato. In questo caso, l'E.Coli per risparmiare energia produce enzimi necessari per digerire il lattosio, solo una volta presente nell'intestino.
Nel 1961 Jacob e Monod formularono l'ipotesi secondo la quale l'E.Coli usa tre enzimi per assorbire e iniziare a metabolizzare il lattosio. I geni che codificano per questi enzimi sono attivati dallo stesso sistema di regolazione e sono vicini tra loro. Accanto a questo gruppo di geni vi sono due sequenze di controllo, che partecipano alla loro regolazione. Una sequenza di nucleotidi corrisponde al promotore, il sito in cui si attacca l'RNA-polimerasi che catalizza la trascrizione. Un segmento di DNA chiamato operatore agisce invece come interruttore tra il promotore e questi tre geni. L'operatore decide quando l'RNA-polimerasi può attaccarsi al promotore e iniziare quindi la trascrizione dei geni. Un simile aggregato è chiamato operone ed è presente solo nei procarioti. L'operone Lac (lattosio=glucosio + galattosio) inizia a operare quando E.Coli viene a contatto col lattosio e tutti gli enzimi necessari alla sua demolizione vengono assemblati contemporaneamente poiché i geni dell'operone sono controllati tutti dall'operatore.
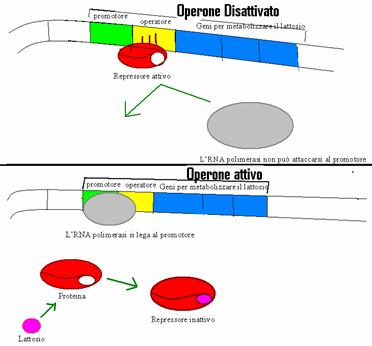 Quando l'operone lac è disattivato,
non è presente lattosio nell'ambiente cellulare. La trascrizione
è bloccata da una molecola chiamata repressore,
una proteina che funziona legandosi all'operatore e impedendo all'RNA polimerasi di attaccarsi
al promotore. Il repressore proviene dal gene
regolatore, situato all'esterno dell'operone.
Quando l'operone lac è disattivato,
non è presente lattosio nell'ambiente cellulare. La trascrizione
è bloccata da una molecola chiamata repressore,
una proteina che funziona legandosi all'operatore e impedendo all'RNA polimerasi di attaccarsi
al promotore. Il repressore proviene dal gene
regolatore, situato all'esterno dell'operone.
Per attivare l'operone lac, è necessario che il lattosio si leghi al repressore cambiandone la forma e impedendo l'attacco all'operatore. È solo così che l'RNA polimerasi può legarsi al promotore e cominciare a scorrere lungo i geni dell'operone.
o Un secondo tipo di operone, detto trp, e controlla la sintesi degli enzimi per la produzione del triptofano(amminoacido). Al contrario del'operone lac, esso è controllato da un repressore che è inattivo quando si trova da solo; per attivarsi deve combinarsi con una piccola molecola specifica.
o Un terzo tipo di operoni usa gli attivatori, proteine che attivano gli operoni legandosi al DNA. Queste facilitano il legame dell'RNA-polimerasi con il promotore, invece che bloccarlo. Con questa varietà di operoni l'E.Coli e gli altri procarioti possono sopravvivere in ambienti mutevoli.
Negli organismi eucaristici, durante le ripetute divisioni cellulari, che porteranno uno zigote a diventare un organismo pluricellulare adulto, le singole cellule vanno incontro al differenziamento, ossia diventano specializzate nella struttura e nelle funzioni. Il differenziamento è il risultato dell'espressione selettiva dei geni, alcuni vengono attivati, mentre altri rimangono inattivi. Ad esempio, le cellule muscolari sono lunghe e dotate di più nuclei. Il pancreas, organo che produce gli ormoni insulina e glucarone per la regolazione della concentrazione del glucosio nel sangue, è diviso in due parti: l'endocrina (produce gli ormoni che vanno nel sangue) e l'esocrina (riversa gli ormoni fuori dal sangue). Qui il gene che codifica per l'insulina, è attivo in alcune cellule, e quello che codifica il glucarone, è attivo in altre cellule.
Le cellule differenziate esprimono solo una piccola percentuale dei loro geni ma per capire se sono ancora potenzialmente in grado di esprimerli, negli anni cinquanta, Steward e i suoi collaboratori, trasferirono alcune cellule di carota in un terreno di coltura; alcune cellule singole e isolate iniziarono a dividersi dando origine ciascuno a una pianta adulta, identica a quella di partenza. Questa tecnica può essere utilizzata per produrre migliaia di organismi geneticamente identici, detti cloni, a partire dalle cellule somatiche di un'unica pianta; ciò dimostra che il differenziamento non comporta cambiamenti irreversibili al DNA delcellula - METABOLISMO, LA RESPIRAZIONE, RESPIRAZIONE AEROBICA DELLA SOSTANZA ORGANICA" class="text">la cellula. Un indizio di questo, è rappresentato dal fenomeno naturale della rigenerazione, ossia la sostituzione di parti corporee perdute che si riscontra in certe specie. Molti animali sono in grado di rigenerare le parti mancanti e a volte, è addirittura possibile che alcune cellule differenziate isolate perdano la loro specializzazione e diano di nuovo origine all'intero organismo.
Se srotolato, il DNA complessivo dei 46 cromosomi di una cellula, raggiungerebbe tre metri di lunghezza. Tutto questo DNA può essere contenuto in un nucleo grande circa 5 mm, grazie a un elaborato sistema di avvolgimento e di ripiegamenti (spiralizzazione). Tale sistema prevede più livelli di complessità ed è caratterizzato dal fatto che il DNA (inizialmente ha 2 nm di diametro) è associato a piccole proteine chiamate istoni. Al primo livello di spiralizzazione gli istoni sono attaccati al DNA. Si vengono così a formare i nucleosomi (10 nm di diametro), formati da un filamento di DNA avvolto due volte attorno a un nucleo proteico formato da 8 istoni. I nucleosomi adiacenti sono uniti tra loro da brevi tratti di DNA, chiamati linker. In un successivo livello di spiralizzazione la collana si avvolge su se stessa, formando una fibra elicoidale compatta (30 nm di diametro). Un ulteriore riavvolgimento di questa fibra dà origine a una struttura elicoidale più spessa con un diametro di circa 300 nm. Successivi livelli di ripiegamento e avvolgimento consentono alla cellula di contenere enormi quantità di DNA. Quindi le cellule possono usare i più alti livelli di spiralizzazione per mantenere a lungo i geni inattivi, visto che per far attivare un gene, gli istoni devono staccarsi dal DNA (despiralizzazione).
Nelle femmine dei mammiferi uno dei due cromosomi X si presenta fortemente condensato in tutte le cellule somatiche e quasi del tutto inattivo. Questa disattivazione del cromosoma X si verifica in uno stadio precoce dello sviluppo embrionale coinvolgendo uno a caso dei due cromosomi X e viene ereditata da tutte le rispettive cellule lie. Ad esempio, i gatti calicot sono riconoscibili per il pelo a macchie arancioni e nere. Il gene calicot si trova sul cromosoma X e il fenotipo richiede la presenza di due differenti alleli: uno per il colore del pelo arancione, e uno non arancione che risulta nero. Se una femmina è eterozigote per il gene calicot, si osserva il fenotipo calicot: le macchie arancioni sono formate da gruppi di cellule in cui è attivo il cromosoma X con l'allele per il colore arancione, mentre le macchie nere sono dovute a cellule in cui è attivo il cromosoma X con l'allele per il pelo non arancione.
Diversamente dai geni degli operoni batterici, ogni gene eucaristico ha di solito una propria serie di sequenze di controllo e in più, sono presenti gli induttori. Lo stato "normale", negli eucarioti pluricellulari sembra essere quello disattivato. Una cellula eucariotica ha bisogno di attivare (quindi di trascrivere) solo quella piccola parte dei geni necessaria per la sua specifica funzione all'interno dell'organismo.
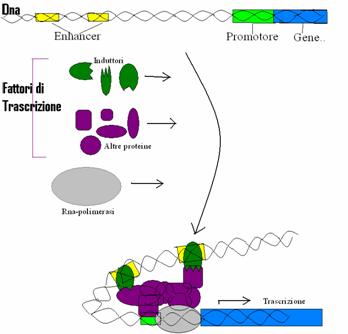 L'attivazione di un gene eucariotico coinvolge, oltre
all'RNA-polimerasi, alcune proteine di regolazione chiamate fattori di trascrizione, tra cui gli
induttori; essi si legano ad alcune sequenze di DNA chiamate enhancer o intensificatori.
Probabilmente il DNA si ripiega e gli induttori interagiscono con altri fattori
di trascrizione che poi si legano tra loro presso il promotore del gene. Questo
complesso di proteine facilita il corretto legame tra l'RNA-polimerasi e il
promotore, e favorisce l'inizio della trascrizione. Oltre agli enhancer e agli
induttori, vi sono anche i repressori, proteine che possono legarsi alle
sequenze di DNA chiamate silencer (silenziatori) e funzionare di conseguenza da
inibitori per l'avvio della trascrizione. In sintesi, sia gli eucarioti sia i
procarioti regolano la trascrizione del DNA tramite proteine che si legano al
DNA, ma nei primi il processo è più complesso.
L'attivazione di un gene eucariotico coinvolge, oltre
all'RNA-polimerasi, alcune proteine di regolazione chiamate fattori di trascrizione, tra cui gli
induttori; essi si legano ad alcune sequenze di DNA chiamate enhancer o intensificatori.
Probabilmente il DNA si ripiega e gli induttori interagiscono con altri fattori
di trascrizione che poi si legano tra loro presso il promotore del gene. Questo
complesso di proteine facilita il corretto legame tra l'RNA-polimerasi e il
promotore, e favorisce l'inizio della trascrizione. Oltre agli enhancer e agli
induttori, vi sono anche i repressori, proteine che possono legarsi alle
sequenze di DNA chiamate silencer (silenziatori) e funzionare di conseguenza da
inibitori per l'avvio della trascrizione. In sintesi, sia gli eucarioti sia i
procarioti regolano la trascrizione del DNA tramite proteine che si legano al
DNA, ma nei primi il processo è più complesso.
Una volta completata la trascrizione, gli introni (segmenti non codificanti), vengono rimossi grazie al processo di splicing. In alcuni casi la cellula può svolgere lo splicing in maniera differente e generare così diverse molecole di mRNA a partire dallo stesso trascritto in RNA. Con questa sorta di splicing alternativo un organismo può ottenere polipeptidi diversi dallo stesso gene.
o Nel citoplasma si trovano degli enzimi che hanno il compito di degradare le molecole di mRNA una volta finita la traduzione. Nei procarioti l'mRNA viene degradato dagli enzimi dopo pochi minuti; questo è uno dei motivi per cui i batteri possono modificare le loro proteine piuttosto velocemente in risposta ai cambiamenti ambientali. L'mRNA degli eucarioti, invece, può sopravvivere diverse ore o perfino settimane.
o I polipeptidi che si formano dopo la traduzione devono essere modificati per diventare funzionali. Negli eucarioti i meccanismi di controllo post-traduzionali contemo spesso l'eliminazione di una porzione del polipeptide. Il risultato è una proteina più piccola e attiva.
o Un altro meccanismo dopo la traduzione è la demolizione selettiva delle proteine. Un tale meccanismo di regolazione permette a una cellula di modificare il tipo e la quantità di proteine in risposta ai cambiamenti dell'ambiente. Inoltre, con questo meccanismo vengono anche eliminate le proteine che hanno accumulato delle anomalie e che devono essere quindi sostituite. L'apparato che controlla questo smantellamento prevede una fase di identificazione delle proteine da degradare, che avviene mediante il legame con piccole sequenze proteiche. Queste, vengono poi riconosciute da un apparato di demolizione detto proteasoma, che le riduce in frammenti più piccoli consumando energia.
La clonazione fornisce numerose prove che le cellule differenziate mantengono appieno il loro potenziale genetico. La clonazione degli animali può essere effettuata tramite il metodo del trasferimento nucleare. Questa tecnica consiste nella rimozione nel nucleo della cellula uovo e nell'inserimento del nucleo di una cellula somatica del donatore adulto nella cellula uovo (priva del suo nucleo). La cellula uovo inizia così a dividersi e, circa cinque giorni più tardi , si forma un embrione allo stadio blastocisti (sfera costituita da circa 200 cellule). Nei mammiferi, dopo questa fase, per ottenere un individuo è necessario impiantare l'embrione nell'utero della madre surrogata. L'animale derivante da questa procedura, detta clonazione riproduttiva, sarà un clone della cellula del donatore (quindi geneticamente identico). Il ricercatore scozzese Wilmut ha usato questa tecnica per clonare la pecora Dolly.
In una differente procedura di clonazione dalla blastocisti si ottengono delle cellule staminali embrionali (cellule ES) che hanno la potenzialità di generare tutte le tipologie di cellule dell'organismo. Queste vengono utilizzate nella clonazione terapeutica, per guarire i pazienti con danni irreversibili a determinati tessuti.
I critici sottolineano che l'attuazione della clonazione umana presenta molti ostacoli, di carattere sia pratico sia etico. Da un punto di vista pratico, la clonazione degli animali è molto difficile e inefficiente. Da un punto di vista etico, la clonazione suscita molti interrogativi. Nel frattempo, la ricerca e il dibattito continuano.
IL CONTROLLO GENETICO DELLO SVILUPO EMBRIONALE
Nelle prime fasi dello sviluppo di un organismo come un moscerino vi sono alcuni geni che stabiliscono a quale estremità della cellula uovo si dovranno formare il capo e la coda.
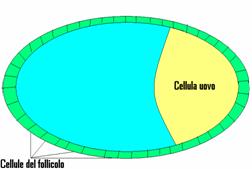 Uno dei primi geni attivati nella cellula uovo codifica per una
proteina che esce da questa e si dirige verso le cellule adiacenti del follicolo.
Uno dei primi geni attivati nella cellula uovo codifica per una
proteina che esce da questa e si dirige verso le cellule adiacenti del follicolo.I ricercatori che hanno studiato i geni omeotici del moscerino della frutta hanno individuato in essi una comune caratteristica strutturale: tutti contenevano una stessa sequenza di 180 nucleotidi. Queste sequenze sono chiamate homeobox. Il segmento polipeptidico dell'homeobox si lega a specifiche sequenze di DNA, consentendo alla proteina omeotica di attivare o disattivare, determinati gruppi di geni.
Per coordinare meglio le attività cellulari in un organismo adulto è fondamentale la comunicazione tra le cellule, un meccanismo con cui certe proteine o altri tipi di molecole portano i messaggi dalle cellule che li trasmettono alle cellule (bersaglio) che li ricevono.
LE BASI GENETICHE DEL CANCRO
Il cancro può essere considerato una malattia nella quale le cellule sfuggono ai meccanismi di controllo che normalmente limitano la loro divisione, ciò avviene perché si modificano i geni di alcune cellule. Uno dei primi passi nella lotta contro il cancro fu la scoperta, nel 1911, di un virus che determina l'insorgenza di un tipo di tumore nei polli. I virus che causano il cancro possono rimanere permanentemente all'interno delle cellule integrando il proprio DNA in quello dell'ospite. Molti virus in grado di provocare il cancro portano nel proprio genoma specifici geni che, una volta all'interno delle cellule ospiti, possono rendere la cellula cancerosa. Un tale gene, che può causare il cancro quando è presente in singola copia nella cellula, è detto oncogène.
Ricerche hanno dimostrato che i cromosomi di diversi animali, compreso l'uomo, contengono proto-oncogèni, cioè geni che possono essere trasformati in oncogèni. Affinché un proto-oncogène divenga un oncogène si deve verificare una mutazione nel DNA, come mutazioni nello stesso proto-oncogène, errori nella duplicazione del DNA o nella ricombinazione, spostamenti del proto-oncogène dal suo sito originario nel DNA.
Il cancro può essere indotto anche da alterazioni dei geni, questi sono detti oncosoppressori perché le proteine che essi codificano contribuiscono a impedire una divisione cellulare incontrollata.
Mutazioni nella sequenza di trasduzione del segnale:
o Nel caso in cui il segnale iniziale sia il fattore di crescita, la risposta finale della cellula bersaglio sarà la sintesi di una proteina che induce la cellula a dividersi. Se questa proteina derivata da un proto-gène chiamato ras, il prodotto normale dovrebbe essere una proteina di trasmissione che, non è in grado di funzionare senza essere sollecitata dal fattore di crescita. La proteina identificata dall'oncogène è invece una versione iperattiva e incontrollata di quella normale.
o Nel caso in cui un fattore inibente la crescita, si leghi con il recettore della cellula bersaglio, si produrrà una proteina che blocca la proliferazione della cellula. L'oncosoppressore p53 codifica per un fattore di trascrizione e inibisce la divisione cellulare. Se avviene un mutamento di questo oncosoppressore tale fattore non inibirà più la divisione cellulare, rendendo possibile una proliferazione anomala.
Mutazioni a carico del gene ras o p53 sono presenti nella maggior parte dei tumori.
Come accade per altri tumori, lo sviluppo del cancro è in grado di produrre metastasi (diffusione oltre il sito di origine), è un processo graduale che in genere impiega una decina di anni per svilupparsi.
Queste modificazioni cellulari sono l'espressione di alterazioni a livello del DNA che avvengono in sequenza, tra cui l'attivazione di un oncogène e l'inattivazione di due geni oncosoppressori. Queste mutazioni genetiche comportano variazioni delle sequenze di trasduzione del segnale. È necessario che si verifichi più di una mutazione (solitamente quattro) nelle cellule somatiche affinché si abbia un completo sviluppo del cancro. Solitamente le prime due mutazioni fanno in modo che le cellule si dividano più rapidamente, apparendo per tutto il resto normali; la terza mutazione incrementa la velocità di divisione cellulare, ma provoca anche cambiamenti nell'aspetto delle cellule stesse; infine, una quarta mutazione cancerogena che determina l'avvio di una divisione incontrollata e l'invasione dei tessuti circostanti.
|
Privacy
|
© ePerTutti.com : tutti i diritti riservati
:::::
Condizioni Generali - Invia - Contatta
