 |
|
| biologia |
|
I VIRUS
Sono microrganismi acellulari, poiché privi delle tipiche strutture cellulari e sono, per tale motivo, dei parassiti obbligati intracellulari.
Sono molto piccoli (30-200 nm) e non possono essere visti al microscopio ottico.
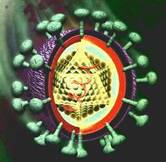 Sono costituiti da un involucro proteico e da un solo acido
nucleico, che può essere alternativamente DNA o RNA.
Sono costituiti da un involucro proteico e da un solo acido
nucleico, che può essere alternativamente DNA o RNA.
In base alla natura dell'involucro, sono da distinguersi i virus nudi dai virus rivestiti; i primi hanno unicamente un involucro di natura proteica chiamato cabside, i secondi oltre al cabside hanno un secondo involucro chiamato pericabside. Fra cabside e pericabside si può poi avere il tegumento.
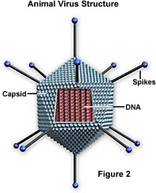
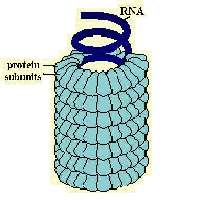
L'involucro può poi essere di diversa forma: si
distinguono in tal senso i virus a simmetria elicoidale dai virus a simmetria
icosaedrica; nei primi le proteine che compongono l'involucro sono disposte
in senso elicoidale e all'interno vi si colloca l'acido nucleico, nei secondi
le proteine si dispongono in modo da formare un icosaedro (solido a 20 facce a
forma di triangolo equilatero), tale disposizione è permessa
dall'associazione di gruppi di cinque proteine (pentoni) e di gruppi di sei proteine (esoni).
L'acido nucleico va a determinare la complessità del virus poiché ne costituisce il genoma: alcuni virus hanno genomi che codificano per poche proteine, altri che codificano per moltissime.
I virus possono essere a DNA ed in tal senso si distinguono molte classi, come quelli con DNA a doppi elica lineare o circolare, quelli con DNA a singola elica. I virus possono poi essere a RNA ed anche in questo caso si distinguono quelli con RNA lineare a singola elica, quelli a RNA a doppia elica, quelli con RNA frammentato.
Esistono poi alcuni virus che durante il loro ciclo replicativi presentano delle forme ibride sia a DNA che a RNA.
Fra i virus ad RNA è poi necessario distinguere fra quelli a polarità negativa e quelli a polarità positiva; i primi hanno un RNA che non funge da messaggero, ma da stampo per gli RNA messaggeri, i secondi hanno un RNA che funziona direttamente come messaggero.
Moltiplicazione dei virus
Il rapporto virus-ospite può essere di natura varia ed in tal senso si distinguono vari tipi di infezioni:
infezione produttiva: l'infezione porta alla formazione di nuove particelle virali;
infezione abortiva: l'infezione non porta alla formazione di nuove particelle, ma la cellula-ospite comunque presenta antigeni virali;
infezione restrittiva: la moltiplicazione del virus avviene solamente in momenti specifici del ciclo cellulare;
infezione persistente: può essere cronica (il virus rimane nella cellula per un lungo periodo e dà un lento e continuo rilascio di nuovi virioni) oppure latente (il virus in alcuni momenti si replica, mentre in altri si sposta in diversi distretti della cellula o dell'organismo per rimanere silente finché, in seguito ad opportuni stimoli, ritorna a moltiplicarsi).
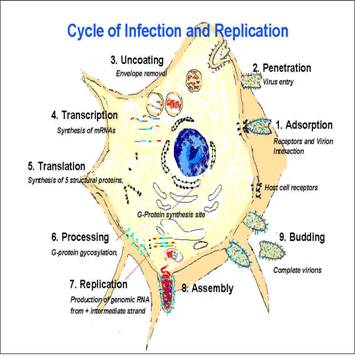 La replicazione
virale, all'interno della cellula, consta di alcuni momenti
fondamentali:
La replicazione
virale, all'interno della cellula, consta di alcuni momenti
fondamentali:
attacco del virus: per mezzo di interazioni recettore-antirecettore, il virus riconosce la propria cellula ospite. A seconda del tipo di recettori che l'antirecettore virale può riconoscere, si va a distinguere lo spettro d'ospite del virus, ovvero il tipo (o i tipi) di cellula che il virus può infettare.
penetrazione del virus: le modalità di penetrazione sono differenti per i virus nudi e per i virus rivestiti:
I virus nudi possono entrare o per endocitosi (attraverso la formazione di una vescicola derivata dalla membrana plasmatica) o per viropepsi (il virus emette degli enzimi specifici che formano un canale nella membrana plasmatica, attraverso cui può essere inoculato il genoma virale).
I virus rivestiti possono entrare per fusione dall'esterno (per mezzo di proteine fusogene il virus aderisce alla cellula) o per fusione dall'interno (il virus viene fagocitato e successivamente, il pH acido della vescicola formata determina la digestione degli involucri virali).
replicazione del virus:
I virus a DNA utilizzano il sistema trascrittivi cellulari, per cui gli enzimi cellulari provvederanno alla trascrizione dell'RNA messaggero e dei nuovi filamenti di DNA, nonché alla sintesi delle proteine virali.
I virus a RNA si distinguono, a seconda della polarità del loro genoma: nel caso di virus a RNA con polarità positiva, l'RNA virale funzionerà immediatamente da messaggero e codificherà per tutte le proteine virali, a cominciare dagli enzimi, sino ad arrivare alle proteine strutturali. I virus a RNA con polarità negativa avranno un genoma che fungerà da stampo per la sintesi di nuovi mRNA; la trascrizione avverrà per mezzo di una RNApolimerasi-RNAdipendente (trascrittasi) di cui il cui il virus è correlata.
assemblaggio e uscita del virus:
Nei virus nudi, le proteine virali strutturali si associano per formare il cabside, entro cui verrà inoculato il genoma virale; in questo modo si formeranno i nucleocabsidi, che usciranno dalla cellula per lisi della membrana.
Nei virus rivestiti, invece, in un primo momento si formerà il nucleocabside; le proteine che costituiranno il pericabside, si legheranno alla membrana plasmatica. A questo punto, si avvicinerà a questo tratto di membrana modificata il nucleocabside e si formerà una vescicola che gemmerà, permettendo l'uscita del virus dalla cellula e l'acquisizione del pericabside.
Le classi replicative
I virus possono essere classificati sulla base del loro genoma, infatti, sulla base di esso, varieranno anche le modalità di replicazione.
Prima classe: virus a DNA a doppio filamento lineare o circolare;
Seconda classe: virus a DNA a singolo filamento (virus dell'herpes labiale);
Terza classe: virus a RNA a doppio filamento;
Quarta classe: virus a RNA a polarità positiva (virus della poliomielite);
Quinta classe: virus a RNA a polarità negativa (virus del morbillo e virus dell'influenza);
Sesta classe: virus a RNA con un intermedio a DNA (virus HIV);
 Settima classe: virus a DNA con un intermedio a RNA (virus
dell'epatite)
Settima classe: virus a DNA con un intermedio a RNA (virus
dell'epatite)
Esempio di prima classe: Herpes simplex (virus dell'herpes labiale)
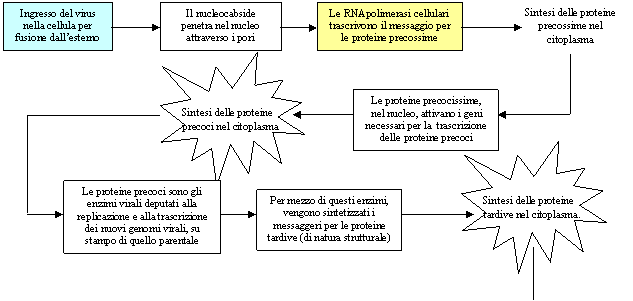
Uscita del virus dalla cellula
![]()


Esempio di quarta classe: Polio Virus (virus della poliomielite)

Esempio di quinta classe
Paramixo Virus (virus del morbillo)
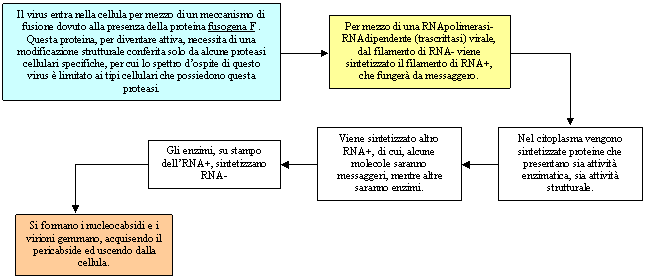
Virus dell'influenza
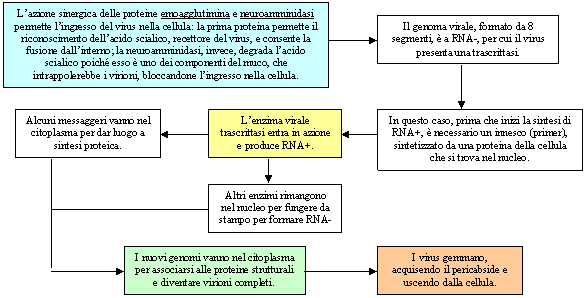
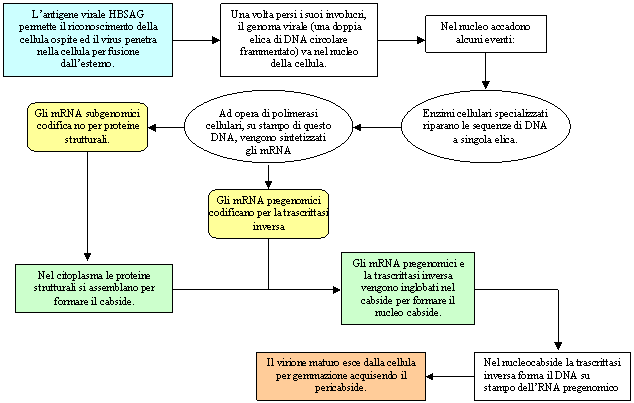
Esempio di sesta classe: Virus HIV

La caratteristica principale dei retrovirus è quella di avere tre sequenze geniche specifiche: GAG, POL, ENV. La prima sequenza codifica per le proteine del cabside, la seconda codifica per gli enzimi virali come la trascrittasi inversa, l'integrasi e una proteasi, la terza codifica per le proteine del pericabside.
HIV possiede poi alcuni geni accessori: il genoma di HIV è definito come diploide dal momento che due sequenze di RNA identiche sono unite all'estremità 5'. Agli estremi del genoma si hanno due geni chiamati rispettivamente U5 e U3 (dove U sta per unico), il primo è localizzato all'estremità 5', mentre il secondo è localizzato all'estremità 3'.
Ai lati di queste sequenze si hanno delle sequenze ripetute (uguali fra loro) chiamate rispettivamente R5 ed R3.
Nel momento in cui si ha la formazione del filamento di DNA a partire da quello di RNA, i geni U5 e U3 si invertono, per cui si avrà il gene U5 all'estremità 3' e il gene U3 all'estremità 5'. Le sequenze ripetute, invece, rimangono uguali fra di loro, per cui in 3' si avrà la sequenza U5OR3, mentre all'estremità 5' si avrà la sequenza U3RO5, le quali sono fra loro identiche.
Queste sequenze, chiamate anche LTR (long terminal repeat) hanno la funzione di regolare il ciclo biologico del virus.
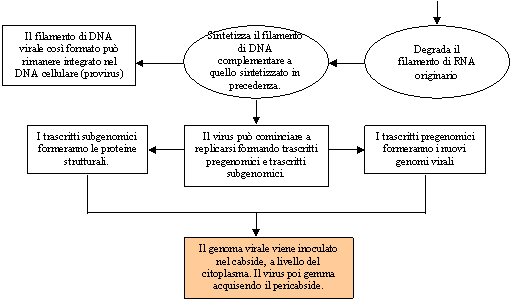
Esempio di sesta classe: Hepatna Virus (virus dell'epatite)
Patogenesi virale
La patogenicità dei virus è conferita dal fatto che i virus sono parassiti endocellulari obbligati ed è data da tutti quegli eventi imputabili alla presenza del virus all'interno della cellula.
Il contatto virus-ospite comincia con l'infezione, la quale poi potrà evolvere, a seconda della natura del virus, in malattia più o meno grave.
Le infezioni possono essere di vario tipo:
Infezione acuta: è un'infezione che dura per brevi periodi di tempo, il decorso è abbastanza rapido e non lascia alcuna complicazione. (un esempio è il virus del raffreddore o dell'influenza).
Infezione persistente
infezione cronica: il virus, in modo lento e costante, rilascia nuovi virioni portando alla cronicizzazione della malattia;
infezione latente: il virus alterna momenti di replicazione a momenti di latenza, se il virus si moltiplica, si hanno i comuni sintomi, mentre nella fase di latenza i sintomi scompaiono per poi eventualmente riire in seguito a particolari stress.
L'infezione è caratterizzata da alcuni eventi:
ingresso del virus nell'ospite: in linea di massima, il contatto virus-ospite avviene a livello delle prime vie respiratorie o a livello del tratto digerente. Nel punto di primo contatto, il virus comincia a moltiplicarsi.
diffusione dal sito di ingresso: alcuni virus possono dare infezioni limitandosi ai punti di ingresso (il virus del raffreddore si limita ad infettare le prime vie respiratorie), altri virus, invece, possono spostarsi a luoghi adiacenti rispetto a quelli della prima moltiplicazione ed altri ancora possono percorrere dei lunghissimi tratti all'interno dell'organismo (ad esempio il virus del morbillo _una delle malattie sistemiche_ entra in contatto con l'ospite a livello delle prime vie respiratorie; da qui passa nel sistema linfatico _generando a livello dei linfonodi una prima risposta immune_ e successivamente al sangue; una volta in circolo, il virus raggiunge midollo osseo, milza e reticolo endoteliale, dove si moltiplica nuovamente; da qui il virus raggiunge poi gli organi bersaglio e compaiono i sintomi tipici del morbillo).
disseminazione agli organi bersaglio
eliminazione del virus: l'eliminazione può avvenire, ancora una volta, o per mezzo delle vie respiratorie o per mezzo del tratto digerente.
Le vie di trasmissione si distinguono in due grandi gruppi cui appartengono diverse sotto-categorie:
via di trasmissione orizzontale: da individuo a individuo
via aerea: le particelle virali possono essere emesse con l'espirazione sottoforma di aerosol.
via oro-fecale: interessa principalmente i virus nudi.
via venerea: l'infezione viene trasmessa con i rapporti sessuali, coadiuvata dalla presenza di numerose colonie batteriche a livello dei genitali.
via cutanea: l'infezione viene trasmessa per mezzo della pelle, sempre coadiuvata dai batteri presenti.
via parenterale: l'infezione viene trasmessa per mezzo del sangue infetto.
Via di trasmissione verticale: da madre a lio
via transplacentare: i virioni vengono mandati al feto per mezzo della placenta; queste infezioni portano a malformazioni fetali o ad aborto.
via perinatale: l'infezione viene trasmessa dalla madre al lio o durante il parto, o durante l'allattamento.
Le difese antivirali ed i farmaci antivirali
Il nostro organismo è capace di rispondere alle infezioni virali con varie modalità differenti.
Si distinguono però delle risposte specifiche e delle risposte aspecifiche:
Le risposte specifiche sono date dalle interazioni fra antigene virale e recettore e dal conseguente riconoscimento; dopo il riconoscimento, infatti, l'organismo attua una risposta specifica sulla base della natura del virus che è entrato in contatto con il nostro organismo.
Le risposte aspecifiche sono invece date da alcune cellule, come i macrofagi e le cellule natural killer, da molecole, come le proteine complemento, che si legano al complesso anticorpi-cellula infettata provocandone la lisi, o a meccanismi indiretti come i processi di infiammazione o l'innalzamento della temperatura corporea.
Anche la cellula è però in grado di dare delle risposte, servendosi di particolari proteine chiamate interferoni. Gli interferoni sono molecole specifiche per il tipo cellulare che le ha prodotte e non per il virus che ha infettato la cellula, per cui anche loro rientrano fra le difese aspecifiche del nostro organismo. Normalmente la loro sintesi è bloccata; l'evento che induce la loro sintesi è l'entrata del virus nella cellula. A questo punto, la cellula infettata, una volta prodotti gli interferoni, inizia a secernerli all'esterno; gli interferoni andranno quindi a legarsi ad altre cellule non infettate, inducendo al loro interno la sintesi di altre proteine inattivate che si attiveranno solo in caso si infezione e andranno a loro volta a bloccare la replicazione virale. Alcune di queste proteine sono la 2'-5'oligoA-sintetasi, che produce delle corte catene di poli-A che andranno ad inattivare gli mRNA virali, oppure una proteino-chinasi che va a fosforillare il fattore d'inizio EF2, responsabile dell'inizio della traduzione dell'mRNA a livello ribosomiale, disattivandolo e bloccando così la sintesi proteica virale.
Gli interferoni possono essere suddivisi in tre gruppi differenti:
interferoni α: sono prodotti da monociti (precursori dei macrofagi), linfociti B e cellule dendriche nel momento in cui queste cellule vengono a contatto con proteine virali o cellule tumorali. Sono proteine non glicosillate che resistono a pH acidi e hanno una buona capacità di diffondersi ed un'azione rapida.
interferoni β: sono prodotti da macrofagi, cellule epiteliali e fibroblasti in risposta alla presenza di acidi nucleici virali, in particolar modo RNA. Sono proteine glicosillate, resistenti a pH acidi, con azione rapida, ma scarsa diffusibilità.
interferoni γ: sono prodotti dai linfociti T in risposta alla presenza di antigeni virali. Sono proteine glicossilate, facilmente denaturabili a pH acidi, con una scarsa diffusibilità ed un'azione lenta, ma prolungata nel tempo. In effetti questi interferoni non svolgono le funzioni tipiche degli interferoni, ma vanno a stimolare il sistema immunitario e la produzione degli altri interferoni.
Per quanto riguarda i farmaci, essi vanno ad agire a livello dei momenti fondamentali della replicazione virale; esistono quindi farmaci che agiscono sul contatto virus-cellula, sull'ingresso del virus nella cellula, sulla trascrizione del genoma virale e sull'uscita del virus dalla cellula.
Farmaci che agiscono sul contatto virus-cellula: questi farmaci agiscono mascherando il recettore cellulare o l'antirecettore virale; un esempio di quanto detto è il Pleconaril, che mima l'antirecettore virale, legandosi ad un'introflessione del pericabside virale ed impedendo il riconoscimento antirecettore-recettore.
 Farmaci che agiscono sull'entrata del virus nella cellula: due farmaci significativi sono l'Amantadina,
per la cura dell'influenza, e il T20,
per la cura dell'AIDS. Il primo farmaco va ad agire sulla proteina M del virus
influenzale che compone il tegumento. Il cambiamento conformazionale di questa
proteina provoca, al momento della fusione dall'interno, il mancato
abbassamento del pH all'interno della vescicola e quindi l'incapacità di
rilasciare all'interno della cellula il genoma virale. Il secondo farmaco,
invece, interagisce con la membrana plasmatica cellulare, che, a questo punto,
non è in grado di indurre il cambiamento conformazionale della GP41, la
proteina fusogena del virus HIV. In questo modo, la proteina rimane inattiva ed
il virus non può entrare all'interno della cellula.
Farmaci che agiscono sull'entrata del virus nella cellula: due farmaci significativi sono l'Amantadina,
per la cura dell'influenza, e il T20,
per la cura dell'AIDS. Il primo farmaco va ad agire sulla proteina M del virus
influenzale che compone il tegumento. Il cambiamento conformazionale di questa
proteina provoca, al momento della fusione dall'interno, il mancato
abbassamento del pH all'interno della vescicola e quindi l'incapacità di
rilasciare all'interno della cellula il genoma virale. Il secondo farmaco,
invece, interagisce con la membrana plasmatica cellulare, che, a questo punto,
non è in grado di indurre il cambiamento conformazionale della GP41, la
proteina fusogena del virus HIV. In questo modo, la proteina rimane inattiva ed
il virus non può entrare all'interno della cellula.
Farmaci che agiscono sulla trascrizione: i farmaci che agiscono a questo livello vanno a bloccare la sintesi proteica virale. Due esempi sono gli interferoni ricombinanti, che hanno comportamento analogo agli interferoni cellulari, ma vengono ottenuti da batteri e miceti ingegnerizzati, e gli oligonucleotidi antisenso, che impediscono la trascrizione.
Farmaci che agiscono sulla sintesi di DNA ed RNA: questi farmaci sono, sostanzialmente,analoghi strutturali dei comuni nucleosidi (guanosina, citosina ecc.) che bloccano la sintesi degli acidi nucleici virali con vari meccanismi: inibendo, per competizione, la polimerasi; legandosi alla catena, ma impedendo l'attacco del nucleotide successivo; sostituendo un nucleotide nella catena completa, ma togliendo il significato all'acido nucleico sintetizzato.
 Alcuni esempi sono la iododeossiuridina,
analogo della timina ed efficace nelle infezioni erpetiche; l'AZT, analogo della timina, la D-deossicitina, analogo della citosina.
Questi farmaci vengono impiegati, in associazione con una proteasi, nella
triplice terapia dell'AIDS, necessaria per scongiurare lo sviluppo di ceppi
resistenti ai farmaci attualmente impiegati.
Alcuni esempi sono la iododeossiuridina,
analogo della timina ed efficace nelle infezioni erpetiche; l'AZT, analogo della timina, la D-deossicitina, analogo della citosina.
Questi farmaci vengono impiegati, in associazione con una proteasi, nella
triplice terapia dell'AIDS, necessaria per scongiurare lo sviluppo di ceppi
resistenti ai farmaci attualmente impiegati.
Si ha poi l'Aciclovir, un analogo della guanosina che risulta essere poco tossico perché selettivo, efficace nelle terapie per la cura dell'Herpes. La selettività è data dal fatto che questa molecola, per diventare attiva, deve essere fosforillata; la fosforillazione è però data da un enzima virale, per cui il farmaco è attivo solamente nelle cellule infettate.
Farmaci che agiscono sull'assemblaggio: molti virus sintetizzano delle poliproteine che, ad opera di enzimi, chiamati proteasi, vengono suddivise nelle singole proteine per mezzo del riconoscimento di specifiche sequenze aminoacidiche. Questi farmaci, come il Saquinovir, imitano queste sequenze e vanno a bloccare la proteasi, impedendo la formazione delle singole proteine e quindi l'assemblaggio dei nuovi virioni.
Farmaci che agiscono sulla neuroamminidasi: esistono dei farmaci anti-influenzali abbastanza efficaci che bloccano la neuroamminidasi. Questo enzima ha infatti il compito di degradare l'acido scialico; essendo quindi analoghi strutturali di quest'acido, tali farmaci vengono riconosciuti dalla neuroamminidasi, ma poi la bloccano limitando l'infezione.
|
Privacy
|
© ePerTutti.com : tutti i diritti riservati
:::::
Condizioni Generali - Invia - Contatta
