 |
|
| biologia |
|
|||||
|
Non ha una ben definita timentazione, le strutture comuni a tutte le specie sono:
→ Parete
→ Membrana plasmatica
→ Massa citoplasmatica
→ Materiale nucleare
Le strutture che invece caratterizzano le diverse specie sono dette accessorie e sono:
→ Capsula
→ Flagelli
→ Pili
→ Spore
PARETE CELLULARE E PEPTIDOGLICANO
E' situata a ridosso della membrana plasmatica verso l'esterno, è una delle caratteristiche specifiche dei procarioti. Il composto che la caratterizza è il peptidoglicano o mureina.
Quest'ultimo è un eteropolimero molto complesso, formato da zuccheri e aminoacidi e sintetizzato esclusivamente dai procarioti. Ogni subunità di peptidoglicano è formata da un parte proteica e una glicanica.
La parte glicanica è costituita da un disaccaride costituito a sua volta da due aminozuccheri:
N-acetil-glucosamina (NAG)
Acido N-acetil- muramico (NAM)
Nel NAG il gruppo ossidrilico dell'atomo di carbonio α è sostituito da un gruppo amminico acetilato. Il NAM deriva dalla N-acetil-D-glucosamina mediante legame etere con l'acido lattico a 3 atomi di carbonio. Questi sono uniti tra loro mediante legami β-l,4-glucosidici. I due aminozuccheri sono comuni nelle diverse specie batteriche.
La parte proteica è costituita da una catena laterale tetrapeptidica, associata al NAM tramite il residuo di acido lattico, gli aa che compongono questa catena sono.
→ L-alanina
→ D-acido glutammico
→ L-lisina nei Gram+ e acido meso-diaminopimelico nei Gram-
→ D-alanina
Caratteristica interessante è la presenza di aa nella conformazione D, rara nelle strutture biologiche e tale da rendere il peptidoglicano resistenti all'azione dei comuni enzimi proteolitici (esoenzimi che degradano le proteine) specifici per gli L-aa.
FASI PRINCIPALI DELLA BIOSINTESI DEL PEPTIDOGLICANO
a) Formazione del nucleotide di Park nel citoplasma, è un precursore che ha anche una D-alanina in posizione 5 .
b) Il nucleotide di Park viene trasferito mediante il bactoprenolo pirofosfato alla zona di crescita del peptidoglicano e durante il trasporto avviene il completamento biosintetico. Questa fase potrebbe essere limitante in quanto il bactoprenolo potrebbe essere bersaglio dell'antibiotico bacitracina che inibisce la fosforilazione del bactoprenolo fosfato, bloccando la sintesi del peptidoglicano.
c) Formazione dei legami covalenti con la porzione glicanica (NAM e NAG) a tra le catene peptidiche (ponti). L'energia per la formazione dei legami deriva dal legame D-alanina in posizione 5 e D-alanina in posizione 4, dato che all'esterno della cellula non esiste ATP per adempiere a tale funzione. Questo fenomeno prende il nome di transpeptidazione.
FUNZIONI DELLA PARETE
♦ Proteggere le cellule dalle sollecitazioni meccaniche
♦ Proteggere le cellule dalla lisi osmotica
Queste funzioni sono state comprese attraverso esperimenti che impegnavano, come agente di demolizione della parete, il lisozima. Quest'ultimo è un enzima che idrolizza il legame β-l,4 glicosidico tra il C1 del NAM e il C4 del NAG. Quando cellule bastoncellari vengono poste in una soluzione isotonica di saccarosio e trattate con lisozima le cellule rimangono vitali ma, a seguito della perdita della parete, diventano di forma sferica e sono chiamate protoplasmi. Le stesse cellule invece, poste in soluzione ipotonica e trattate con lisonzima, muoiono per lisi osmotica. Il peptidoglicano della parete, essendo una struttura rigida che si oppone all'espansione, regola la pressione esercitata dall'acqua dall'interno contro la parete, impedendo un eccessivo rigonfiamento della cellula e una conseguente morte.
Non è sufficiente che ci sia un certo numero di strati per donare resistenza alla parete ma è fondamentale che questi formino una serie di ponti tra aa di anelli diversi. I ponti o legami interpeptidici sono molto importanti ed esistono degli batteri diventano immuni agli antibiotici" class="text">antibiotici specifici, appartenenti alla categoria "analoghi strutturali", come la dicicloserina, che riescono a non far formare questi ponti e quindi a destabilizzare la parete.
Ci sono essenzialmente due tipi di legami interpeptidici:
Nei Gram- , come
E.Coli, in genere il legame avviene tra il m-DAP in posizione 3 di una catena e
Nei Gram+, come
S.aureus, invece il legame interpeptidico è in genere "indiretto" , infatti avviene sempre
tra l'aa in posizione 3 (generalmente una L-lisina) e
Alcuni batteri nel corso dell'evoluzione hanno tratto un grande vantaggio nel poter formare i legami interpeptidici tramite questi ponti. Infatti proprio S.aureus, che è un patogeno, ha evoluto la caratteristica di modificare spessissimo il tipo di ponte che si forma fra le catene di peptidoglicano. Questo, soprattutto per un patogeno, è un'enorme vantaggio in quanto variando il tipo di aa del ponte possono maggiormente difendersi dalla eventuale digestione proteolitica (proteasi).
I diversi studi, condotti a partire dagli anni '50 hanno evidenziato l'esistenza di due modelli strutturali di parete, confermando e spiegando il diverso comportamento dei batteri alla Colorazione di Gram. Questa tecnica prende il nome dal suo ideatore. Tale tecnica utilizza in sequenza due diversi tipi di coloranti:
1ª fase → si usa il cristal violetto per 2-3 minuti, questo colorante colora il citoplasma di tutte le cellule di
color viola, in quanto è in grado di entrare in qualsiasi tipo di cellula.
2ª fase → le cellule sono trattate con una soluzione che contiene iodio. L' unione di iodio e cristal violetto
fa complessate le cellule, cioè laddove c'è unione si forma un complesso più grande che non è
più in grado di uscire dalla cellula. Si fissa in questo modo il colore.
3ª fase → si tratta con un decolorante ( alcol etilico o acetone)
4ª fase → si usa un secondo colorante, la safranina rossa. A questo punto avviene la classificazione:
Gram + , conservano la colorazione viola
Gram - , perdono la colorazione viola e si colorano con la safranina in rosso.
Questo perchè la cellula si disidrata con l'alcol, ciò provoca un compattamento del peptidoglicano:
♦ Se lo strato è spesso → il colorante non riesce a superare l'impedimento (peptidoglicano
compattato) Gram+
♦ Se lo strato è esile → il trattamento con alcol provoca il rilascio del colorante, Gram-
|
GRAM+ |
GRAM- |
|
Si colorano di viola |
Si colorano di rosso |
|
Il 3° aa è L-lisina |
Il 3° aa è l' acido meso-diaminopimelico |
|
I legami interpeptidici sono di tipo diretto |
I legami interpeptidici sono di tipo indiretto |
|
La loro parete appare come uno strato unico. continuo e spesso. |
La loro parete appare pluristratificati e sottile |
|
Sulla parete ci sono anche: acidi teicoici e lipoteicoici che fanno da collante tra il peptidoglicano e la membrana. |
Sulla parete ci sono anche: lipoproteina di Brown o mureina, che fa da ponte tra il peptidoglicano e la membrana esterna(modalità vedi disegno sotto) |
|
Proteine di superficie che creano dei canali per permettere il passaggio di alcune sostanze. |
Porine che di solito si trovano in gruppi di tre, attraverso le quali entrano o escono sostanze |
|
Queste molecole sono vitali per i batteri e se modificandosi con un meccanismo di variazione genica di superficie non intaccano la loro funzione principale e quindi la vitalità della cellula, si modificano assumendo funzione antigenica. |
LPS che l' antigene per eccellenza. E' costituito da 3 regioni, andando dall'interno verso l'esterno: lipide a, ha attività pirogena e tossica core polisaccaridico, costituito da zuccheri parte O, la sua lunghezza non è bene definita in quanto è costituita da un'unità strutturale ripetuta n volte. E' la parte che funge da antigene, più i patogeni saranno efficienti quanto più questa porzione somiglierà alle glicoproteine umane. |
|
Es. virus dell'influenza, del morbillo e della rosolia. |
Es Artrite reumatica, ceppo K4 di E. coli ha la condroitina |
CAPSULA
Alcuni batteri possono avere esternamente alla membrana esterna anche una capsula, che nella maggior parte dei casi è di natura polisaccaridica o a volte anche proteica.
Fa parte delle strutture accessorie e identifica la patogenicità di un batterio. Ha 3 funzioni fondamentali:
Consente al patogeno di aderire al substrato (tessuto o mucosa su cui agire). Proprio per questo motivo, viste le proprietà chimico-fisiche degli zuccheri, essa è in genere polisaccaridica (molto appiccicosa).
Nasconde gli antigeni al sistema immunitario, quindi un batterio capsulato non viene facilmente individuato.
L'ultimo aspetto importante
è che un antibiotico che contro un batterio non capsulato avrebbe
facilità ad entrare nel citoplasma, con la capsula questo ingresso
diventa più difficoltoso, quindi bisogna elevare
FUNZIONI DELLA MEMBRANA
♦ Contiene citoplasma;
♦ Barriera semipermeabile, seleziona l'ingresso e l'uscita delle molecole;
♦ Produzione di energia, inglobano glucosio tramite energia solare. In assenza di glucosio la maggior
parte dei batteri è in grado di sintetizzare composti alternativi come galattosio e lattosio. Attivando
dei geni che codificano per essi che fino a quel momento si trovavano in stato OFF.
♦ Trasporto, che può essere di tre tipi:
. Trasporto semplice → Simporto
→ Antiporto
→ Uniporto
. Traslocazione di gruppo, è specifico e potrebbe accogliere più di una molecola
. Sistema di trasporto ABC, riguarda il dominio proteico che impiega ATP. Da un lato c'è
il carrier che accomna la molecola al trasportatore e dall'altro c'è la proteina che
idrolizza ATP fornendo energia al trasportatore. Da entrambi i lati ci sono proteine
accessorie che incrementano la specificità di passaggio in quanto più il passaggio è
specifico più sarà utile alla cellula.
♦ Motilità, può avvenire mediante delle proteine di membrana che consumano ATP e si legano al
substrato modificando la propria conformazione e quindi i batteri si muovo per scivolamento.
Oppure mediante flagelli che si trovano sulla superficie della membrana. I flagelli possono trovarsi
ad una sola estremità, ad entrambe le estremità oppure su tutta la superficie. Non sono dotati di
motilità propria ma ricevono l'impulso dal corpo basale. In assenza di nutrienti i batteri compiono
dei movimenti orari e antiorario come per perlustrare la zona. Nel momento in cui percepiscono il
cambiamento di concentrazione del nutriente tendono ad avvicinarsi ma se non è a loro gradito si
allontano. Questo fenomeno è detto chemiotassi.
E' importante saper come si divide una cellula batterica e come si risale ad una carica batterica, se per esempio si vuol fare un'analisi di un campione di gamberi per conoscere il numero preciso di organismi batterici e quindi determinare se quella data partita di prodotto è idonea al consumo umano, bisogna determinare, appunto, la carica batterica.
Procediamo per gradi.
Nei batteri la
divisione cellulare, che avviene essenzialmente con un meccanismo di ciccione
binaria, coincide con la riproduzione. 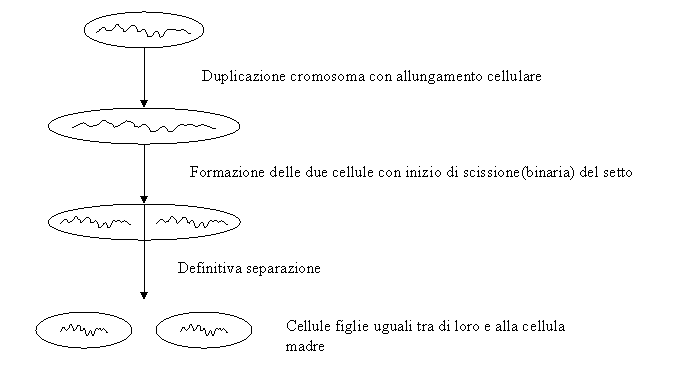

Spesso per alcune cellule, lo step limitante è la replicazione del cromosoma, nel senso che il tempo di duplicazione cellulare, dipende dal tempo che il cromosoma ci mette a replicarsi.
La pericolosità di alcuni ceppi batterici dipende spesso proprio dal tempo di duplicazione. Quelli che velocemente si duplicano, penetreranno meglio e più facilmente causeranno malattie.
Il setto può essere:
♦ Asimmetrico → se è spostato in prossimità di uno dei due poli della cellula di partenza. In questo caso
si parlerà di sporulazione, cioè meccanismo attraverso il quale si forma una spora
batterica.
♦ Simmetrico → si trova in posizione centrale e si avrà la divisione cellulare.
Facendo una sezione di una cellula batterica in divisione ciò che è importante notare è una proteina detta FTSZ. Tale proteina si colloca al centro a livello del setto, crea un anello intorno alla zona di divisione e senza di essa la duplicazione non avverrebbe.
In alcuni casi non si ha una divisione totale, cioè le cellule lie al termine della divisione rimangono attaccate dando origine ad aggregati di cellule. L'esempio con maggiore varietà di disposizione è dato dai cocchi che si possono presentare come:
♦ Diplococchi, si dividono lungo un piano e rimangono attaccate a due a due;
♦ Streptococchi, si dividono lungo lo stesso piano e rimangono attaccate a formare delle catene;
♦ Stafilococchi, si dividono su + piani, in modo irregolare, rimanendo attaccate a formare dei grappoli;
♦ Sarcine, si dividono lungo i tre piani, in modo regolare formando aggregati geometrici di tipo cubico.
Trattazione matematica della divisione
Una cellula batterica, trovandosi in condizioni ambientali favorevoli, origina due cellule che, a loro volta, si dividono in due, per un totale di quattro cellule, le quali, riproducendosi, portano il numero delle cellule a otto, e così via. Quindi ad ogni generazione corrisponde un numero di individui pari al doppio di quello della generazione immediatamente precedente. In tal modo, partendo da un singolo individuo, si ottiene un insieme di individui, cioè una popolazione.
Quindi l'aumento del numero delle cellule conura una tipica progressione geometrica:
1→ 2→ 4 → 8 → 16 → . → 2*2 ( n-l)
20 → 21 → 22 → 24 → . → 2n
L'esponente indica il numero di generazioni.
L'intervallo di tempo impiegato da 1 o N cellule per diventare 2N, quindi per raddoppiare è detto tempo di generazione.
Un altro parametro importante è la velocità di crescita cioè il numero di generazioni nell'unità di tempo.
Esercizio tipo nel calcolo del numero di cellule
Abbiamo in una beuta
un terreno con 3*103 cellule in 30ml di soluzione. Il ceppo
batterico ha come tempo di generazione
Ci saranno 4 generazioni:
3*103cellule : 30ml = 100 cellule in 1 ml
100*24 = 1600cellule/ml (dopo
Oppure:
3*103*24 = 48000
cellule in 30ml (dopo
4.8*104 : 30 = 1600cellule in 1ml (dopo
Come standard si usa sempre 1ml
Ritmo di crescita
Non è ipotizzabile che i batteri continuino a raddoppiare sempre di numero, altrimenti dopo qualche giorno avremo una massa batterica equivalente a quella di un transatlantico. Naturalmente ad un certo punto c'è una limitazione alla crescita batterica che può essere causata da:
♦ esaurimento della fonte esogena di energia;
♦ mancanza di nutrienti essenziali;
♦ presenza di prodotti metabolici che inibiscono la crescita.
Curva di crescita
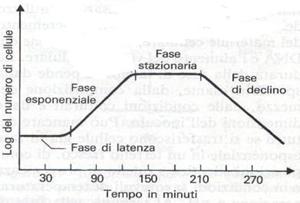
♦ Fase di latenza o fase lag, durante la quale i batteri, pur disponendo di abbondanza di nutrienti o non si
replicano per niente o si accrescono lentamente. Rappresenta infatti la fase di adattamento
alle nuove condizioni ambientali e sono pertanto in piena attività metabolica.
♦ Fase esponenziale o fase log(logaritmica), caratterizzata da una velocità di crescita costante che
dipende dalla specie, dal mezzo e dalle condizioni di coltura. Le cellule presentano lo
stesso tempo di generazione, anche se non tutte si dividono esattamente nello stesso
momento. Infatti ad ogni istante nella coltura, sono presenti cellule in tutti gli stati del
ciclo vitale. Se il mezzo di coltura non viene rinnovato, dopo alcune generazioni, la
crescita esponenziale si interrompe e si passa alla fase stazionaria.
♦ Fase stazionaria, caratterizzata da una velocità di crescita nulla, dovuta all'arresto della moltiplicazione
cellulare o a una forma di compensazione tra il tasso di riproduzione e quello di mortalità.
I fattori che causano tale fase sono quelli detti sopra. Tale fase segna il passaggio da una
popolazione giovane a una popolazione fisiologicamente vecchia. Con l'età anche il
comportamento rispetto alle colorazioni può variare. I batteri gram+ possono diventare in
questa fase, gram variabili o gram-, per l'incapacità di mantenere la parete in buone
condizioni.
♦ Fase di declino o di morte, caratterizzata da una diminuzione di tipo esponenziale del numero di cellule
vive, per cui ad ogni intervallo di tempo, la metà delle cellule sopravvissute muore. Ciò è
dovuto al prevalere nella coltura di condizioni particolarmente sfavorevoli.
Se prendiamo una popolazione batterica in piena fase esponenziale e la trasferiamo in un terreno ricco di nutrienti, vedremo che immediatamente i batteri riprenderanno a replicarsi secondo il loro tempo di generazione.
Se invece trasferiamo in questo terreno batteri in fase stazionaria o in fase di morte, essi avranno bisogno di un certo tempo per riadeguare il proprio apparato proteico, biosintetico e catabolico, alla disponibilità di nutrienti e quindi in realtà risultano essere in fase di latenza.
Nella fase stazionaria i batteri percepiscono di trovarsi in una situazione di pericolo, perché non riescono più a nutrirsi e a seconda della specie adottano dei meccanismi di adattamento.
Le specie sporigene, laddove scarseggiano i nutrienti e si intraprende la fase stazionaria, si dividono asimmetricamente formando le spore, in modo da non aver più bisogno di nutrienti. Le spore infatti sono una forma di criptobiosi, ovvero la vita è nascosta nella spora stessa. Il DNA viene protetto in un involucro e non c'è bisogno di energia, perché tutto resta spento. La spora quindi è una forma di resistenza alla condizione avversa di mancanza di nutrienti. Gli sporigeni, i bacilli, i clorigeni, gli streptoliceti, "decidono" ( in effetti tutto è sempre basato su una reazione e fenomeni biochimici) di sporulare e quindi proteggere il DNA. Quando ci sarà nuovamente disponibilità di nutrienti, l'involucro viene eliminato e il DNA verrà di nuovo trascritto ed espresso e quindi il batterio "esce" dalla forma vegetativa e ricomincia il ciclo batterico vitale.
Ci sono altri adattamenti come gli Attinomiceti, i quali competono per la sopravvivenza.
Una determinata nicchia ecologica, presenta varie specie batteriche. Se si è in fase stazionaria, questi batteri competono con gli avversari, producendo molecole capaci di uccidere altri batteri. Ecco come si sono evoluti i primi antibiotici.
Queste specie batteriche hanno iniziato a produrre, in fase stazionaria, quei metabolici secondari che non sono altro che antibiotici capaci di bloccare la crescita o uccidere altre specie batteriche, liberandosi così dell'avversario.
Ovviamente il produttore di un antibiotico deve essere resistente all'antibiotico, altrimenti sarebbe un suicidio.
Paradossalmente, i produttori di antibiotici che devono essere resistenti all'antibiotico e hanno un gene che gli conferisce quella resistenza, sono anche la causa principale della diffusione di quella resistenza. Tale gene si è infatti "diffuso" in specie diverse da quelle produttrici.
FABBISOGNO ENERGETICO
Le due forme di energia che possono essere utilizzate dal procarioti sono: l'energia radiante della luce solare e l'energia chimica delle molecole nutritive. In entrambe i casi l'energia viene impiegata per sintetizzare ATP. Gli organismi capaci di catturare l'energia solare si dicono fototrofi o fotosintetici e sono: piante verdi, alghe, i cianobetteri e alcuni altri gruppi batterici. La fonte di energia chimica è costituita dalla presenza nell'ambiente di composti chimici che, una volta introdotti nelle cellule, cedono energia attraverso reazioni di ossidazioni. Gli organismi che operano queste trasformazioni si dicono chemiotrofi o chemiosintetici. Oltre agli animali sono chemiotrofi i funghi, i protozoi e la maggior parte dei batteri. L'ossidazione dei substrati nutritivi viene effettuata dai microrganismi secondo due processi fondamentali: la fermentazione e la respirazione. La fermentazione è tipica dei lieviti e di molti batteri, è costituita da una serie di reazioni che avvengono in condizioni anaerobiche, cioè in assenza di ossigeno molecolare, e che comportano la degradazione di un substrato nutritivo, generalmente un carboidrato, in un composto organico a minor contenuto energetico. Il principale prodotto finale definisce il tipo di fermentazione:acida, lattica, acetica ecc.
La respirazione, simile come processo biochimico in tutti gli organismi viventi, presenta nei batteri alcune peculiarità. Alla più comune respirazione aerobica, in cui all'ossidazione del substrato organico si accomna la riduzione dell'ossigeno molecolare, si affianca la respirazione anaerobica che utilizza, come accettori finali di elettroni, sostanze inorganiche diverse dall'ossigeno, quali nitrato, solfato, carbonato. I microrganismi che dipendono esclusivamente dalla respirazione aerobica sono aerobi obbligati. I microrganismi che vengono inibiti dall'ossigeno molecolare e impiegano processi fermentativi o ossidazioni anerobiche si definiscono anaerobi obbligati. Mentre i microrganismi capaci di crescere sia in assenza sia in presenza di ossigeno si chiamano anaerobi facoltativi.
FATTORI AMBIENTALI
CHE INFLUENZANO
Per ogni specie microbica si possono individuare dei valori minimi, ottimali e massimi di temperatura, entro cui avviene la crescita.
♦ Temperatura
minima, è
♦
Temperatura ottimale, è
♦ Temperatura massima, limite superiore entro cui la crescita è ancora possibile; oltre tale valore
intervengono fenomeni di denaturazione proteica comportanti non solo il blocco della crescita ,ma
anche la morte cellulare.
In base ai valori delle tre temperature i microrganismi vengono classificati in tre categorie:
♦ Se l'optimum è abbastanza elevato parliamo di batteri termofili.
♦ Se l'optimum è più o meno temperatura ambiente parliamo di batteri mesofili.
♦ Se l'optimum è a temperatura bassa di batteri psicrofili.
Esistono dei meccanismi di adattamento per mantenere la temperatura interna costante:
♦ Esistono degli enzimi detti desaturasi e saturasi che inseriscono o eliminano legami multipli nell'acido
grasso. In particolare a basse temperature la membrana tende a gelidificarsi impedendo il nomale
svolgimento delle attività biologiche, alcuni batteri psicrofili hanno acidi grassi particolari, poco
ordinati, ciò è utile in quanto inserendo doppi legami nella catena idrocarburica un acido grasso saturo
diventa insaturo (vengono aggiunti doppi legami). Si impedirà così al batterio di gelificarsi troppo.
Ad alte temperature, saranno acidi grassi saturi, senza doppi legami a "proteggere" il batterio.
♦ Chaperonine, proteine da shock termico che sono capaci di tenere la proteina distesa, nel caso in cui va
in contro a denaturazione per le alte temperature.
♦ CPS, proteine da shock freddo. In quanto a basse temperature l'mRNA può strutturarsi in maniera
secondaria, ovvero si formano parziali zone a doppia elica. Le CPS si legano all'mRNA impedendo la
formazione delle strutture secondarie. Man mano scorre il ribosoma e le CPS vengono così liberate. La
loro funzione è quella di tenere l'mRNA lineare.
Prendiamo in considerazione il parametro pH. Troveremo batteri che riescono a vivere in condizioni di pH estreme. Alcuni riusciranno a sopportare acidità di 1, altri acidità 13.
In realtà il batterio non tiene il suo valore di pH uguale a quello esterno, bensì riesce a tenere il suo citoplasma sempre (+ o -) neutro, nonostante ambienti acidi o alcalini. Vivono in ambienti estremi mantenendo il proprio pH + o - uguale.
In genere i batteri si riproducono asessualmente e si scambiano materiale genetico mediante 3 modalità:
♦ Trasformazione
♦ Trasduzione
♦ Coniugazione
TRASFORMAZIONE
E' un processo mediante il quale i batteri assumono DNA presente in forma solubile nell'ambiente. E' un processo versatile in quanto non esiste una grossa specificità tra le specie batteriche tra cui avviene la trasformazione.
Il DNA esterno prende contatto con la superficie batterica dove incontra una nucleasi che degrada una delle due eliche. L'elica integra entra nel citoplasma dove incontra le SSBP che legano il singolo filamento prevenendo il riappaiamento. Nel citosol c'è anche un'altra proteina detta RECA che favorisce la ricombinazione con il cromosoma integrandolo in esso. A questo punto avviene la duplicazione in modo tale che il DNA esterno vada a far parte del corredo genomico. Il batterio ricombinate avrà a questo punto migliori capacità di adattamento rispetto a quello non ricombinato in quanto assorbendo DNA dall'esterno ha acquisito tutte le proprietà di questo e donerà alla progenie il o i geni acquisiti.
Le SSBP sono delle proteine che hanno dei batteri cosiddetti competenti come streptococcus e stafilococcus, un batterio diventa competente nel passaggio di transizione tra la fase esponenziale e stazionaria perché se termina il nutrimento in quell'ambiente, inglobando un pezzo si DNA estraneo può capitare che potrà assimilare anche componenti presenti nell'ambiente stesso.
Cerca così di integrare geni per utilizzare altri substrati, traendo vantaggio e quindi poter continuare o riprendere la fase esponenziale.
Questo fenomeno è tra l'altro usato in processi di creazione e ingegnerizzazione di cellule batteriche.
TRASDUZIONE
E' il processo mediante il quale il DNA passa da una cellula donatrice ad una ricevente mediante un fago(virus batterico). Il faco è specie specifico quindi il passaggio avverrà tra specie uguali o simili.
Il
fago ricoperto del capside contatta la cellula batterica immettendo al suo
interno il DNA fagico, lasciando all'esterno il capside. Il DNA fagico diventa
il substrato per l'RNApolimerasi che inizia a trascrivere, il primo gene che
trascrivere codifica per una proteina endonucleasica essenziale per il fago in
quanto spezzetta il cromosoma del batterio. A questo punto
I capsidi che contengono il genoma batterico, inizialmente infetteranno perché è il capside l'agente infettante. Però tale infezione non si svilupperà perché nel momento in cui il DNA batterico verrà trasdotto non è capace di infettare. Verrà integrato se l'ospite lo riterrà opportuno.
Nell'ambito del ciclo infettivo dei virus può accadere che il virus anziché seguire la via della lisi segue la via lisogenica ovvero il genoma del virus che ha infettato la cellula batterica anziché distruggere il suo genoma si integra in esso replicandosi. Diventando così un profago ovvero un precursore del fago. Però se le cellule si trova in condizioni di stress il profago percepisce questo pericolo passa dalla lisogenia alla lisi e fuoriesce dal cromosoma batterico.
Nell'operazione di fuoriuscita può avvenire una ricombinazione difettosa a causa della quale si forma un genoma virale non perfettamente integro e completo ma con parziale genoma batterico. In questo caso quando andrà ad infettare una nuova cellula, l'infezione non sarà completa quindi si ha comunque una trasduzione ma prenderà il nome di trasduzione specializzata in quanto ad essere trasferito sarà il pezzo di DNA in cui si era inserito il profago. Quindi l'infezione non sarà completa ma la trasduzione del Dna batterico nel batterio ospite si.
ANTIBIOTICI
Sono molecole di origine biologica prodotti da alcuni microrganismi, sono tossici per i batteri ma non dannose per le cellule ospiti, sono quindi dei farmaci a tossicità selettiva per i soli microrganismi procarioti dove interferiscono con il metabolismo e la riproduzione cellulare, determinando azione batteriostatica o battericida. L'abuso degli antibiotici ha portato alla frequente sa di ceppi batterici antibiotico-resistenti. La resistenza agli antibiotici è frutto delle mutazioni spontanee casuali che si verificano in tutte le specie batteriche. Se la mutazione è cromosomica, il gene mutato può essere trasferito ad altri batteri della stessa specie per mezzo della coniugazione o per mezzo di fagi. In questo modo si ha la trasmissione della resistenza verso un solo antibiotico. Se invece i geni responsabili della antibiotico-resistenza sono localizzati in sede extracromosomica (plasmidi) e vengono trasferiti per coniugazione o per mezzo di fagi si può verificare il trasferimento di plasmidi portatori di più caratteri di antibiotico-resistenza che possono diffondersi inoltre anche ad altre specie batteriche.
BATTERI SPORIGENI
Diversi batteri sono in grado di produrre delle forme cellulari particolari, specializzate per sopravvivere in condizioni non favorevoli. Queste forme cellulari di resistenza si chiamano spore e poiché si originano sempre all'interno della cellula madre sono più precisamente indicate con il termine endospore. La cellula normale, senza spora, prende il nome di cellula vegetativa, mentre la cellula portatrice della spora si chiama sporangio. Le spore rappresentano uno stato di vita quiescente: sono metabolicamente quasi inattive, resistono alle alte temperature, all'essiccamento, alle radiazioni ultraviolette, ad agenti chimici nocivi come acidi e disinfettanti, alla carenza di nutrienti e all'invecchiamento. La principale funzione è dunque quella di proteggere il menome da condizioni ambientali sfavorevoli, attraverso un involucro isolante e resistente.
STERILIZZAZIONE
I processi in grado di distruggere i batteri e gli altri microrganismi nell'ambiente esterno sono molteplici:
♦ Sterilizzazione, è un processo ad ampio spettro, diretto alla distruzione di tutte le forme di vita
microbiche. In microbiologia, più esattamente nei laboratori viene impiegata la sterilizzazione in
autoclave che si basa sul vapore sotto pressione. Infatti la sterilizzazione viene effettuata alla
temperature di
una buona sterilizzazione in autoclave deve essere allontanata l'aria presente, in quanto se il
vapore è misto all'aria la conduzione del calore è rallentata.. Eseguito il trattamento di
sterilizzazione, l'apertura dell'autoclave deve essere effettuata solo quando la pressione interna è
scesa ai valori della pressione atmosferica.
♦ Ebollizione,
basato sulla temperatura di
sterilizzazione, in quanto non distrugge le spore batteriche. La sterilizzazione con calore umido a
vapore fluente utilizzando l'autoclave ma evitando l'aumento di pressione.
♦ Tindalizzazione, è un metodo di sterilizzazione frazionata. A un primo trattamento termico che uccide le
forme vegetative, viene fatto seguire un periodo d'incubazione di 24 ore che favorisce la
germinazione delle spore. Il
materiale così trattato viene riportato alla temperatura di 80-
30 minuti, al fine di uccidere le cellule vegetative originatesi dalla germinazione delle spore.
Queste operazioni vanno ripetute due o tre volte. Viene utilizzata per sostanze che non sopportano
le alte temperature.
♦ Pastorizzazione,
è un trattamento a temperature inferiori ai
l'eliminazione della maggior parte dei microrganismi e soprattutto dei germi patogeni, compresi i
virus. Viene utilizzata soprattutto nel settore alimentare proprio per uccidere eventuali germi
patogeni e per ridurre la carica microbica di alimenti.
♦ Calore secco, si realizza in stufe a circolazione di aria calda operando, per eliminare spore e forme
vegetative, a temperature di 160-
contenitori di vetro o oggetti metallici.
TERRENI DI COLTURA
Per poter coltivare i batteri in laboratorio occorre utilizzare degli idonei mezzi di coltura detti terreni di coltura. Sono costituiti da materiale nutritivi in grado di riprodurre artificialmente, in laboratorio, l'ambiente che può soddisfare le esigenze metaboliche del batterio. In base al loro stato fisico i terreni vengono distinti in liquidi e solidi. Per poter solidificare un terreno liquido occorre aggiungere l'agar un polisaccaride acido estratto di alghe rosse, che non risulta tossico per i batteri e non viene attaccato enzimaticamente. Il terreno di coltura liquido o brodo di coltura è fondamentale in batteriologia ed è la base per altri terreni più specifici. Il terreno solido e liquido sono considerati anche terreni di base in quanto a seconda dell'aggiunta di altre sostanze prendono il nome di:
♦ Terreno di arricchimento, al brodo normale vengono aggiunti sangue, siero ecc in modo tale da favorire
lo sviluppo dei batteri eterotrofi particolarmente esigenti che si vogliono isolare rispetto ai
batteri contaminati.
♦ Terreni selettivi, sono resi tali con l'aggiunta di opportune sostanze in grado di svolgere azione di
batteriostasi verso tutti batteri, fatta eccezione per quelli che si vuole coltivare, che viene quindi
favorito nello sviluppo.
♦ Terreni differenziali, sono resi tali dall'aggiunta di sostanze e indicatori per es di pH in grado di rivelare
peculiari attività metaboliche che si verificano nel terreno in seguito alla crescita della specie
batterica in esame.
 |  |
|
Privacy
|
© ePerTutti.com : tutti i diritti riservati
:::::
Condizioni Generali - Invia - Contatta
