MICROBIOLOGIA - BATTERIOLOGIA
INTRODUZIONE
La microbiologia è la scienza che studia i più piccoli organismi
viventi: i microbi (da micron, piccolo, bios, vita, logos, scienza). Questa
scienza nasce con le scoperte di Luis Pasteur, che si distacca dalla teoria
della generazione spontanea, considerato come dogmatica dalla chiesa, e
impostando un nuovo pensiero. La teoria della generazione spontanea, è
la teoria secondo la quale la vita si può originare dall'inanimato: i
topi, secondo questa, nascono da vecchie camicie e stracci; le mosche dalla
carne marcia. Furono gli studi di Francesco Redi, sulle larve degli insetti, di
Lazzaro Spallanzani e di Luis Pasteur a far cadere queste convinzioni. Nella
ricerca microbiologa è importante l'opera di Antony Van Leewenhock, un
commerciante di stoffe di
Delft che nel 1650 fu il primo a vedere materialmente i microbi, tramite un
microscopio di sua invenzione, ma il segreto di tale strumento morì con
lui, ritardando di molto il progresso scientifico. Luis Pasteur, tramite
l'esperimento delle bottiglie di cigno, dimostrò che la teoria della
generazione spontanea, identificò, inoltre, i responsabili della
fermentazione (i lieviti) utilizzati nel campo gastronomico ma non conosciuti,
e i responsabili della decomposizione dei cibi; sviluppò la tecnica
della pastorizzazione, portò prove sostegno delle malattie infettive
sviluppando la teoria del germe, secondo la quale le malattie infettive
originano da germi e non da aria inquinata, sviluppò vaccini contro la
rabbia e altre malattie.
Nel 1910 Paul Ehrlich, sviluppando l'idea di un proiettile magico, che colpisse
selettivamente l'agente patogeno, senza aggredire l'ospite; quest'idea si
basava sul fatto che alcuni coloranti coloravano selettivamente solo i batteri
ma non le cellule umane. Egli così scoprì il salvarsan, composto
di sintesi (chemioterapico) contenente l'arsenico, che dimostrò la sua
efficacia contro Treponema pallidum, agente eziologico della sifilide. Nel 1928
Alexander Flemming scopre casualmente la penicillina, derivato dalla
Pennicillinae notatum, che si dimostrò il primo farmaco antibatterico
specifico. Nel 1945 il microbiologo statunitense Selman Worksman scopre la
streptomicina, sostanza prodotta naturalmente da Streptomyces griseus
(antibiotico). Negli anni successivi furono introdotti molti vaccini come:
vaccino antipolio di Salk (IPV), vaccino antipolio di Sabin (OPV), morbillo,
orecchioni, rosolia, epatite B.
KOCH E I POSTULATI
Robert Koch, famoso scienziato tedesco vissuto tra il 1843 e il1910, che
identificò l'agente eziologico dell'antrace e della tubercolosi (che da
lui è chiamato bacillo di Koch), introdusse tecniche di cultura pura e
ancor più importante introdusse i postulati che da lui prendono il nome.
I postulati di Koch, se sono rispettati da un microrganismo, lo identificano
come agente della malattia infettiva in esame; questi enunciano: il germe deve
essere presente nell'organismo nel corso della malattia, deve essere isolato in
forma pura, gli isolati devono dare la stessa malattia se reinoculati in
animali sensibili, il germe deve essere reisolabile dalla nuova vittima.
PROCARIOTI, EUCARIOTI E VIRUS
Dal punto di vista morfologico i microbi possono dividersi in: eucarioti,
procarioti e acellulari o virus.
Gli eucarioti hanno un nucleo ben definito, in quanto delimitato dalla membrana
nucleare, il DNA al suo interno è organizzato in cromosomi, con
patrimonio genetico diploide ossia con geni costituiti da due alleli con le
quali combinazioni si creano condizioni di omozigosi eterozigoti. Nel
citoplasma è presente un complesso sistema di organuli che comprende: i
mitocondri, i cloroplasti, i lisosomi, i centrioli, complessi sistemi vacuolari
che includono il reticolo endoplasmtico liscio e rugoso e l'apparato del Golgi.
Le strutture più superficiali comprendono la semplice membrana cellulare
o complesse strutture rigide di cellulosa. I movimenti e affidato a movimenti
citoplasmatici con andamento ameboide. La cellula eucariotica ha grandezza
superiore a 5µm; fanno parte degli organismi eucariotici i protozoi e funghi.
I microbi procarioti sono caratterizzati da cellule con dimensioni tra i 1 µm e
i 5 µm; molto semplici, queste cellule hanno un unico filamento circolare
cromosomico che, sprovvisto di membrana nucleare, è immerso nel citoplasma.
Non sono presenti organuli e la capacità di movimenti e data da
flagelli. I microbi procarioti comprendono i batteri classificato secondo: la
forma, la dimensione, la colorazione al Gram, la motilità, la presenza
della capsula, la capacità di sporificare, la capacità di
utilizzare ossigeno, la morfologia della colonia, il DNA e attraverso anticorpi
commerciali (Streptococcus pneumonite= Cocco, Gram positivo, immobile,
asporigeno, capsulato, colonie a punta di spillo).
I virus sono costituiti da informazione genetica affidata a un acido nucleico,
costituito da RNA o DNA a doppio o singolo filamento, protetto da una struttura
proteica chiamata capside, e rivestito o no da un involucro lipidico detto
peplos. Privi di citoplasma non sono considerati viventi, visto che al di fuori
del cellula ospite sono inerti e privi di metabolismo, e non hanno la
capacità di muoversi. La grandezza dei virus va da 30nm a150 nm
(1nm=10-9 m =10-3 µm). La classificazione si basa su forma e dimensioni sulla
presenza del peplos, e sulla natura del genoma.
IL MICROSCOPIO OTTICO ED ELETTRONICO
Un osservatore normale, ad occhio nudo, ha come limite di risoluzione, ossia la
distanza più piccola fra due oggetti che consente di vederli come
separati, di 0.2mm, al disotto di queste grandezze necessitano strumenti
appositi. Il microscopio ottico, che si basa su una serie di lenti che
permettono ingrandimenti successivi, ha limite di risoluzione di 0.2µm e
permette di studiare cellule eucariotiche, e procariotiche. Il microscopio elettronico,
che è molto più complesso, e creato nel 1931 ci ha permesso lo
studio di virus; il suo limite di risoluzione è 0.5nm.
COLORAZIONI GRAM E ZIEHL-NEELSEN, PREPARATI A FRESCO
Cellule eucariotiche e procariotiche si studiano con il microscopio tramite
preparazioni a fresco o preparati fissati e colorati.
La preparazione a fresco viene fatta con cellule vive in sospensione in una
goccia d'acqua, tale goccia sarà posta fra o due vetrini, o in
un'incavatura in un vetrino (cellula di Koch) e coperta da un vetrino copri
oggetti, oppure in una cella in rilievo fra due vetrini. Spesso nella tecnica a
fresco si utilizzano coloranti come inchiostro di china (metodo Burri), che
conferisce una colorazione negativa che ci permette di riconoscere varie strutture.
I metodi più in uso utilizzano preparati colorati e fissati; queste
tecniche fanno uso di germi morti, lo scopo della colorazione e di rendere
visibili strutture interne ed esterne del batterio. Le metodologie di
colorazioni più usate sono: la colorazione Gram, e la colorazione
Ziehl-Neelsen (colorazioni differenziali). La colorazione Gram viene fatta
strisciando su di un vetrino i batteri e fissati al calore, a questo punto si
aggiunge cristalvioletto mordenzato (i mordenti sono sostanze che favoriscono
la fissazione dei coloranti) con acido fenico, si lava alcool etilico 80°, si
aggiunge una soluzione di iodo-iodurata (liquido di Lugol) per un minuto, si
lava e si aggiunge acetone-etanolo per trenta secondi, e infine si aggiunge
safronina per un minuto. I batteri che si colorano di viola vengono detti Gram
positivi, quelli che si colorano di rosa vengono detti Gram negativi, tale
differenza sta nella diversa struttura della parete. Alcuni hanno comportamento
incostante con la colorazione Gram e sono detti Gram variabili. La colorazione
di Ziehl-Neelsen viene fatta strisciando su di un vetrino i batteri, si
aggiunge carbofucsina e si flamba per un minuto, si lava e si aggiunge
iodo-iodurata, si aggiunge una soluzione alcool-acido e si aggiunge blu di metile.
La colorazione Ziehl-Neelsen è utile nei batteri Gram variabili.
CRITERI DI CLASSIFICAZIONE
Oltre alla colorazione Gram la forma è un altro criterio di
classificazione per lo studio di batteri, le forme principali sono tre:
sferico, a bastoncino e a spirale.
I cocchi hanno forma sferica; essi possono presentarsi come singoli elementi e
sono detti micrococchi, o raggruppati in due o in più elementi, tali
raggruppamenti rappresentano i piani spaziali in cui si compie la divisione
cellulare. Alcuni elementi si riuniscono due a due, questi sono chiamati
diplococchi; specie che si raggruppano a forma di grappoli sono detti
staphylococchi, la divisione di questi avviene in due o in tre piani; cocchi
che si dividono in un piano e tendono ad assumer la forma di catene sono detti
streptococchi; le sarcine assumono forme cubiche dividendosi in due o tre
piani.
I bacilli hanno forma a bastoncino la cui forma varia molto tra specie e
specie: alcuni sono più arrotondati e vengono detti coccobacilli, altri
hanno i lati paralleli e le estremità nette; solitamente, la forma nella
stessa specie resta uguale, mentre lunghezza e larghezza possono variare molto.
Alcuni baccilli tendono a disporsi l'uno vicino all'altro parallelamente con un
movimento detto slipping, altri con un movimento detto snapping si dispongono
ad angolo acuto formando V (microbatteri).
I batteri a forma a spirale comprendono numerose varianti, le più
importanti sono: i vibrioni con forma di bastoncini ricurvi a virgola, i
spirilli con forma a spirale, e le spirochete con curve ravvicinate.
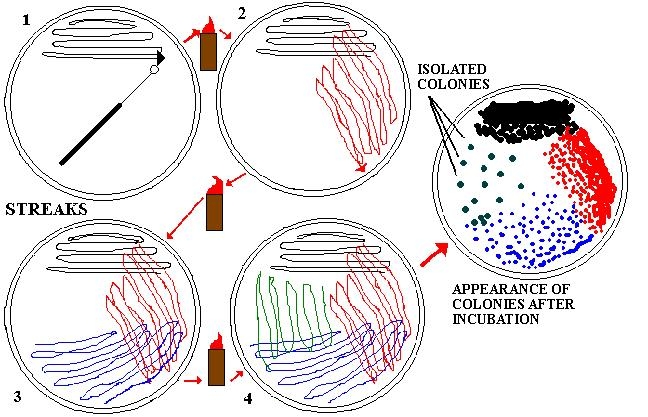
La colonia batterica è un'insieme di cellule lie che derivano da
un'unica cellula genitrice cresciute dopo almeno 24 ore in terreni solidi,
quindi se si ottiene una colonia si è isolato un'unica specie batterica.
La morfologia della colonia è un criterio di classificazione, infatti,
possiamo dividere le colonie per forma di cui le distinguiamo in puntiformi,
tonde, fusiformi, irregolari, irregolari; secondo i margini che possono essere
interi, lobato, filamentoso, lamellare, ondulato e secondo i profili che sono
piatti, elevati, convessi, concavi, umbonati.
LE PRICIPALI FAMIGLIE PATOGENE PER L'UOMO
Secondo queste classificazioni dividiamo i batteri in :Cocchi Gram-positivi,
aerobi come Staphylococcus (S. aureus) e Streptococcus (Str. pyogenes); Cocchi
Gram-positivi, anaerobi come Peptostreptococcus; Cocchi Gram-negativi, aerobi
come Neisseria (N. meningitidis, N. gonorrhoeae); Cocchi Gram-negativi,
anaerobi come Veillonella; Bacilli Gram-positivi, aerobi come Bacillus, (B.
anthracis, C. dyphtheriae); Bacilli Gram-positivi, anaerobi come Clostridium
(Cl. tetani, Cl. perfringens) che forma spore e Lactobacillus e
Propionibacterium; Bacilli Gram-negativi aerobi come Enterobatteri (Escherichia
coli) Haemophilus; Bastoncelli ricurvi (Vibrio cholerae, Campylobacter,
Helicobacter); Micobatteri M. tuberculosis, Treponema (T. pallidum) non
coltivabile; Mycoplasmi privi di parete, intracellulari obbligati, come
Rickettsia, Chlamydia, Coxiella.
LA STRUTTURA DEI BATTERI
I procarioti unicellulari, ossia i batteri, hanno dimensioni tra i 0.2µm e i
10µm; sono studiabili al microscopio ottico, ma l'avvento del microscopio
elettronico ha aumentato le nostre conoscenze sulla struttura cellulare. Le
strutture di una cellula batterica si dividono in essenziali e facoltative: le
prime sono il nucleoide, la membrana cellulare e la parete batterica; le
seconde sono i plasmidi, il mesosoma, la capsula, la membrana esterna, i
flagelli e infine i pili o fimbrie.
La cellula batterica è sprovvista di un nucleo circondato dalla membrana
nucleare, al suo posto ha il nucleoide, costituito da un unico cromosoma
circolare, che comprende, per Eschelicia coli, circa 5*106 basi, e da un fitto
intreccio di fibrille.
Oltre al DNA cromosomico, i batteri possono possiedere DNA extracromosomico,
ossia i plasmidi; questi generalmente portano geni non essenziali per la
riproduzione del batterio, ma contengono geni che codificano per la resistenza
ai farmaci. Molti di questi plasmidi possono essere trasferiti da un batterio a
un altro mediante un sistema di scambio del materiale genetico, detto
coniugazione.
Il citoplasma è quanto mai semplice, mancano gli organuli e il sistema
del fuso, sono invece presenti molti ribosomi, che liberi nel citoplasma gli
danno un aspetto granulare, inoltre rappresentano il 40% del peso cellulare. I
ribosomi batterici sono diversi da quelli eucariotici, costituiti da due
subunità, la 50S e la 30S. Inoltre nel citoplasma sono contenuti granuli
metacroatici (che si colorano di rosso con blu toluina) caratterizzati da
accumuli di fosforo, granuli di glucosio e zolfo.
La membrana cellulare ha la tipica struttura bilaminare come quella
eucariotica, costituita di proteine, lipidi e glucidi nella forma di
glicoproteine e glucolipidi. I lipidi per la maggior parte sono fosfolipidi,
mentre non c'è presenza di steroli a differenza della membrana
eucaristica. La composizione della membrana tra Gram negativi e Gram positivi
è diversa. Le funzioni della membrana cellulare sono: un controllo
selettivo dei trasporti in entrata e in uscita dalla cellulae inoltre svolge
molte funzioni enzimatiche ( come alcune di quelle che negli eucarioti vengono
svolte dagli organuli) come la sintesi della parete. La membrana non è
continua, ma presenta alcune invaginazionicome i mesosomi, che svolgono un
importante ruolo nella divisione dei nuclei (il cromosoma si ancora li durante
la replicazione batterica), o strutture fotosintetiche.
La parete batterica, propria solo dei batteri, permette al batterio vita anche
in condizioni estreme, protegge la membrana impedendo la lisi; essa è
ance responsabile della forma ed anche dell'attacco di molti antibiotici. La
parete è formata da peptidoglicano, o mureina, un enteropolimero
biologico formato da tre parti: uno scheletro di natura polissaridica, da cui
si dipartono corte catene polipeptidiche e da legami crociati fra le catene
peptidiche. Lo scheletro polisaccaridico è formato da due zuccheri:
l'N-acetilglucosamina (o NAG), e l'N-acetilmuramato (NAM), legati fra loro in
sequenza alternata con legami ?1,4 gluosidici (che vengono rotti dal lisozoma).
Le catene tetrapeptidiche sono legate al gruppo carbossilico del NAM, ed sono
formate da L-alanina, D-acido glutammico, da un gruppo R variabilee da
D-alanna; il gruppo R può essere fatto di L-lisina, o meso-L-acido
diamminopimelico (DAP), o L-DAP, o L-ornitina, o L-acido diamminobutirrico
oppure L-omoserino. I legami crociati variano tra le specie batteriche,
comunque nella maggior parte si istaurano legami tra il terzo e il quarto
amminoacido di una catena successiva. I Gram positivi si differenziano dai Gram
negativi da una maggior quantità di pepticoglicano e da una maggior
quantità di legami crociti; nei Gram positivi, inoltre, tutti i NAM fanno
legami con i tetrapeptidi, mentre nei Gram negativi solo uno ogni due o tre o
dieci. I spessori calcolati al microscopio elettronico, dimostrano che nei Gram
positivi sono presenti più strati di mureina, a differenza dei Gram
negativi che ne possiedono uno. Una maggiore quantità di legame
crociati, una maggior quantità di mureina a strati, e la presenza di
acidi tecoici fanno della parete dei Gram positivi più resistente di
quella dei Gram negativi. Gli acidi tecoici sono composti ricchi di fosforo che
si dividono in: acidi glicotecoici, che si legano alla membrana citoplasmatica,
e acidi ribitoltecoici, che si proiettano nella parte esterna della parete; la
funzione degli acidi tecoici è di dare maggiore resistenza. La sintesi
del pepticoglicano è un complesso processo che avviene in tre fasi:
nella prima i precursori dell' enteropolimero vengono sintetizzati nel
citoplasma, nella seconda vengono trasportati al di fuori della membrana da un
trasportatore liposolubile, nell'ultima vengono agganciati fra loro a opera di
enzimi della membrana.
Nei Gram negativi è presente un'altra struttura ancora più
esterna: la membrana esterna. Tra membrana esterna e la parete è
presente lo spazio periplasmico, che occupa il 20%-40% del volume totale; essa
contiene svariate proteine di trasporto, enzimi litici e degradanti che
proteggono la cellula da antibiotici, e oligosaccaridi derivati dalla membrana
(MDO) che hanno il compito di proteggere la cellula dall'iperosmolarità
e dall'iposmoralità. La membrana esterna ha la struttura di una tipica
membrana biologica: è formata da due strati, quello interno
fosfolipidico, mentre quello esterno costituito da lipopolisaccaridi(LPS); la
sua funzione è protettiva , impedisce l'entrata di sostanze dannose
(come la penicillina) ma permette l'uscita di cataboliti tramite proteine di
trasporto. Il lipopolisaccaride è definito come una endotossina, in
quanto non viene liberato se non con la lisi cellulare. La sua struttura di
base è uguale a tutti i Gram negativi: un polisaccaride, e un lipide. Il
lipide viene detto lipide A, esso è custode delle proprietà
tossiche che mantiene anche separato dal polisaccaride, che è invece
responsabile delle specificità antigeniche. Il lipide A è formato
da un acido grasso a lunga catena legato a un disaccaride di diglucosammina
difosfato; a questo si legano due porzioni: un polisaccaride core e un
polisaccaride O. Il polisaccaride CORE (nucleo) è comune a tutta la
specie, ed costituito da 7-8 zuccheri, mentre il polisaccaride O è
proprio del ceppo. La produzione del lipopolisaccaride è compito della
membrana citoplasmatica, che unisce un acido grasso a un al disaccaride di
diglucosammina difosfato, i polisaccaride CORE viene aggiunto attaccando
l'acido 2cheto-3-desossictulonico (KDO) al lipide A e trasportato attraverso i
siti di Bayer punti di continuità tra la membrana esterna e quella
cellulare; infine il polisaccaride O, ripetizione di 40 unità, viene
aggiunto dall'esterno in blocco. Le proteine presenti sulla membrana esterna
sono le porine, formate da quattro subunità(OmpC OmpA OmpF PhoE), che si
uniscono a formare canali che permettono l'entrata di soluti, e da
lipoproteine, la più abbondante, unisce covalentemente la membrana
esterna alla mureina garantendo stabilità alla struttura. LPS è
causa di gran parte dei sintomi della infezioni da Gram negativi provocando
effetti diretti e indiretti. Gli effetti diretti sono l'attivazione della
coagulazione, e del complemento: la coagulazione della coagulazione comporta la
formazione di coaguli nei capillari, e in particolari casi, se per esempio
avvengono nel surrene possono causare la morte, mentre nei casi migliori
causano petecchie; l'attivazione a cascata delle proteine del complemento,
normalmente presenti nel sangue, causa buchi nella membrana cellulare. Gli
effetti indiretti sono immunomediati, come lo shock endotossico e la febbre:
quando un macrofago si lega all'LPS, precedentemente legato a una proteina,
vengono liberate dal macrofago sostanze che causano l'infiammazione come le
interleuchine, prostaglandine e citochine, che causano la febbre, ma se la
produzione è eccessiva viene causato lo shock endotossico, che
può portare alla morte.
La capsula è un criterio di classificazione del ceppo, essa è la
conseguenza del riversamento verso l'esterno della parete cellulare di grossi
polimeri che costituiscono uno strato mucoso superficiale molto sviluppato. La
capsula non ha funzione vitale ma ha funzione antifagocitaria, lo studio di
questa è resa possibile con l'inchiostro di china, che non entra nella
capsula e costituisce una colorazione negativa.
I flagelli sono strutture filamentose responsabili della capacità di
muoversi del batterio: questi ruotano sia in senso orario che antiorario
rimanendo rigidi, spingono la cellula batterica in ogni direzione. La parte
rotante viene detta corpo basale, che inserita nella membrana e collegata a
polimeri di flaggellina che formano un uncino; il corpo basale , è stato
calcolato che può compiere da 10 a 20 giri al secondo.
I pili o fimbrie sono strutture più piccole dei flagelli atte
all'adesione o allo scambio di materiali, che si dividono in pili comuni e
sessuali. I pili sessuali sono geneticamente causati da plasmidi, che
può essere scambiato attraverso questi da un donatore F+ a un ricevente
F-, che a sua volta diventerà F+; a differenza dei pili sessuali, quelli
comuni sono più piccoli e numerosi, i pili sessuali inoltre presentano
cavità centrali e rigonfiamento terminale; entrambi sono costituiti di
polipeptide formato da subunità di pilina.
I PIÙ IMPORTANTI BATTERI GRAM NEGATAVI
I più importanti batteri Gram negativi sono: della Famiglia delle
Enterobatteriaceae come Salmonella che causa diarrea, setticemia e febbre,
Shigella che causa dissenteria, la Klebsiella pneumoniae che causa Polmoniti,
Eschelichia coli Proteus che causa affezioni delle vie urinarie; la famiglia
delle Neisseriaceae come Neisserea meningitidis che causa meningite epidermica,
Neisserea gonorrhoeae, che causa gonorrea e altre famiglie come Haemophylus che
causa Otite e meningite, Brucella che causa febbre maltese, Pseudomonas che
causa infezioni nosocomiali, Bordetella che causa pertosse e Vibrio colerae che
causa il colera.
LE SPORE
Alcuni batteri Gram positivi appartenenti al genere Baccilus, Costridium e
Sporosarcina, in condizioni sfavorevoli alla vita, hanno la capacità di
formare spore. Le spore batteriche sono endospore che si formano attraverso un
complesso processo di differenziazione cellulare, per cui la cellula vegetativa
da origine al suo interno a una cellula differente in struttura e composizione.
Le spore sono resistenti al calore, all'essiccamento, all'azione di
disinfettanti, alle radiazioni, agli antibiotici e possono resistere fino a
milioni di anni. Queste stupefacenti caratteristiche di resistenza sono dovuti
al fatto che sono metabolicamente dormienti, inoltre sono in uno stato a secco;
se l'ambiente sarà di nuovo favorevoli, tramite un processo detto
grerminazione, darà di nuovo in vita alla forma vegetativo. Eistono
inoltre esospore batteriche, non di grande importanza, e inoltre spore fungine
, che non hanno nulla a che fare con quelle batteriche.
La struttura della spora matura è una disposizione degli elementi a
molti strati. La parte centrale viene detto protoplasma o nucleo centrale, che
costituito dal DNA, dal RNA e da proteine. Esternamente al protoplasma
c'è la membrana interna, che deriva dalla membrana cellulare, e al
difuori di questa c'è uno strato di pepticagliacano detto corteccia; fra
questi due c'è uno strato elettrondenso di peptidoglicano che
formerà la parete della cellula vegetativa durante la germinazione.
Superiormente alla corteccia un rivestimento esterno fatto di un materiale
simile alla cheratina, a volte sopra di questa si trova una struttura esterna
detta esosporio di natura lipidica.
Finita la vita vegetativa comincia il processo di sporulazione, che avviene in
diverse tappe: il DNA viene replicato è la cellula divide
asimmetricamente (settazione asimmetrica), formando una prespora, la cellula
vegetativa comincia a ingolfare la prespora producendo la corteccia di
pepticoglicano e la disidrata, si forma anche il rivestimento esterno della
spora, fatto di proteine simili alla cheratina,e in alcuni casi si forma anche
l'Esosporio lipidico esterno; la spora viene liberata e la madre si lisa e
muore. La germinazione è il processo inverso alla sporulazione, sembra
che quando l'ambiente ritorni favorevole, la spore tramite recettori lo
percepisce e germina. I sporigeni più importanti sono: i Clostridium
tetani, difficililis e botolinum, e il bacillus anthracis.
LA REPLICAZIONE BATTERICA
La replicazione batterica avviene per scissione binaria: il DNA comincia a
essere duplicato in un punto detto replicatore, che nell'estremità 3' si
fissa al mesosoma, aquesto punto comincia lo rotolamento per rotazione facilitato
dall'allontanamento del mesosoma, l'etremità 5' si lega alla membrana e
mentre il mesosoma si allontana i filamenti vengono duplicati. I due punti in
cui i cromosomi sono legati sono separato da un setto traverso dove, una volta
terminata la duplicazione del DNA, comincia la citochinesi. Questo processo
viene detto semiconservativo in quanto nelle due cellule che si originano,
metà di entrambi DNA deriveranno dalla cellula materna.
I TERRENI
Una specie microbica può essere conoscere compiutamente solo dal momento
in cui viene isolata in una coltura pura, intendendo per coltura pura lo
sviluppo di una determinata specie microbica ben definita. La premessa di ogni
cultura pura, è la disponibilità di un terreno di cultura nel
quale siano contenute tutte le necessità della specie microbica in
studio, e tale da consentire operazioni fondamentali quali l'isolamento e lo
sviluppo. I terreni possono essere liquidi, dei quali il più comune
è il più comune è quello fatto di brodo di carne, che
permettevi avere grandi quantità di batteri già isolati; esistono
terreni a base vegetale o con lieviti arricchiti con siero, bile, o zuccheri.
Al fine di isolare culture pure però sono indispensabili terreni solidi.
Questi terreni solidi vengono fatti aggiungendo agar a un terreno liquido;
l'agar è un sostanza estratta dalle alghe rosse del genere genere
gelidum composta per il 70% da agarosio e per il 30% da agaropectina. La
propieta del agar è quella di fondere a 100° C, e intorno a 40° di
solidificare in una massa gelatinosa. Il brodo viene aggiunto all'agar fuso, le
cellule batteriche vengono intrappolate con le gocce del terreno liquido nelle
maglie polisaccaridiche dell'agar, che non permette loro di spostarsi,
ottenendo culture pure. I terreni solidi a base di agar, vengono chiusi in tubi
o in scatole Petri, inoltre possono essere aggiunti diversi substrati come il
sangue (per lo più di pecora), e in questo caso si parla di agar sangue.
Un terreno viene detto selettivo se permette l'accrescimento preferenziale di
solo certe specie patogene, attraverso l'azione di sostanze tossiche selettive,
inattive sulla specie di interesse, ma attive sulle altre. Per esempio terreni
di sale mannite permettono la crescita a staphilococcus ma non a streptococcus;
il terreno di Mc Conkey, contiene Sali biliari che permettono la crescita a
enterobatteri, ma letali ad altri batteri. Un terreno viene detto differenziale
se è presente un indicatore che ci permette di riconoscere le differenti
specie batteriche: il terreno di sale mannite è anche un terreno
differenziale, in quanto si colora di diversi colori le differenti specie, per
esempio staphilococcus aureus si colora di giallo. Ai fini della crescita del
germe, il terreno deve possedere alcune caratteristiche: deve contenere forme
di energia (sostanze organiche per i chemioeterotrofi) o la possibilità
dell'utilizzo della luce, composti chimici del carbonio e dell'azoto, per la
sintesi degli amminoacidi, inoltre, deve contenere accettori di idrogeno quali
solfiti, nitriti e carbonati per gli anaerobi, mentre ossigeno per gli
anaerobi, e infine amminoacidi, vitamine, fosforo, zolfo e metalli.
I fattori ambientali sono quegli agenti importanti per la crescita di un
batterio, essi sono: la temperatura, il pH, la presenza di ossigeno e
l'osmolarità. La temperatura è un fattore molto importante,
alcuni batteri detti psicrofili vivono tra -5°C e i 30°C, altri, detti
termofili vivono tra i 25°C e gli 80°C; i batteri di interesse medico sono i
mesofili, questi batteri vivono tra i 35°C e i 40°C. I valori ottimali di pH
per i batteri è presso la neutralità, tra i 7 e i 7.5; questo
però è molto influenzato dai cataboliti, il loro aumento porta
alla morte. L'osmolarità non è un fattore molto importante in
quanto la parete e l'MDO anche per grandi variazioni protegge la cellula.
Alcuni germi detti alofili necessitano della luce solare come Staphilococchi.
L'ossigeno risulta essenziale per alcuni, velenoso per altri, e indifferenti
per altri ancora. Secondo ciò distinguano aerobi obbligati che vivono
solo in presenza di ossigeno, anaerobi obbligati, per cui l'ossigeno è
tossico (Clostridium tetani), i microaerofili che richiedono poco ossigeno,
aerobi facoltativi e aeroindifferenti che possono vivere sia con che senza.
Tenendo conto di ciò un terreno per anaerobi viene incubato sotto vuoto
o con atmosfere di azoto o monossido di carbonio.
CONTA DEI BATTERI E CURVA DI CRESCITA
La conta dei batteri in una cultura risulta molto importante per valutare il
metabolismo, la virulenza e l'efficacia di un antibiotico; la conta può
essere fatta per massa microbica o per numero di germi. Per valutare la massa
batterica i metodi più usati sono: il peso secco, pesando i precipitati,
oppure per densità ottica della cultura liquidi tramite densitometri che
emettono fasci di luce che tramite la diffrazione di questi calcolano la
torpidità della cultura. Per valutare il numero di germi può
essere fatto o per conta al microscopio elettronico utilizzando camere di conta
appositamente fatte, oppure diluendo serialmente, piastrando, e infine contando
le colonie: partendo dalla sospensione di studio si diluisce serialmente fino a
110 di ml, a questo punto si piastra e si incuba per 24 ore a 37°C; infine si
contano le colonie formatesi e si moltiplica per il reciproco della diluizione
(considerando che una colonia sia la discendente di un'unica cellula), a questo
punto ho le CFU, ossia le unità capaci di formare colonie.
La curva di accrescimento dei batteri studia l'andamento di una popolazione in
condizioni standard, utilizzando le più semplici metodiche di conta. La
curva viene fatta inoculando in un terreno liquido dei batteri, mescolando
periodicamente e prelevando piccoli campioni a istanti prefissati calcolo il
numero di germi tramite uno spettrometro; raccolti i dati disegno una curva
riportando sull'ascissa i tempi e in ordinata il numero di batteri. Se si
utilizza una carta semi logaritmica riportando i tempi in progressione
logaritmica posso distinguere quattro fasi: la fase di latenza dovuta
all'adattamento al terreno, in cui i batteri producono enzimi; la fase
esponenziale o logaritmica, dove la velocità di divisione è
massima, seguendo la progressione geometrica; la fase stazionaria in cui tanti
ne nascono e tanti ne muoiono, dovuta all'alta presenza di cataboliti, la fase
di morte dove tutte muoiono. Risulta chiaro la curva è diversa da
cellula a cellula (Eschelichia coli muore in 24 ore).
LA FERMENTAZIONE
Con il termine fermentazione batterica si intendono una serie di reazioni
anarobe che hanno come primo reagente il piruvato derivato dalla glicolisi, il
quale scopo è la riduzione del NADH in NAD. I vari tipi di fermentazione
sono: l'alcolica, la lattica (streptococcus), lattica mista(batteri lattici),
la butilen glicolica, la butirrica, acetone butanolica(clostridium) e infine
propionica (Propiobatteri, usata nell'industria casearia). Il tipo di
fermentazione è importante per definire la specie, in quanto ogni specie
ha la sua particolare; tramite il test di metil rosso possiamo riconoscerla.
DISINFEZIONE E STERILIZZAZIONE
La disinfezione è un processo in cui l'applicazione di un agente
disinfettante causa la morte dei soli organismi patogeni e si differenzia dalla
sterilizzazione che conduce all'uccisione di tutti i microrganismi. Sia la
disinfezione che la sterilizzazione sono riferiti a oggetti e sostanze. Con il
suffisso -cida si indica un agente capace di uccidere un dato agente, con il
suffisso -statico si indica un agente che ferma la replicazione ma una volta
tolto questa ricomincia. La lotta ai germi viene influenziata da vari fattori
quali il numero totale dei germi presenti, dal terreno, dalla natura dei germi,
dall'agente utilizzato e dal tempo di esposizione, dalla presenza di materiali
estranei (nutrienti) e dal pH. Il metodo di sterilizzazione più
importante è il calore, che trova largo impiego anche in ambiente
ospedaliero. Il calore agisce sui batteri coagulando e inattivando le proteine,
o fondendo i lipidi delle membrane; il calore può operare in presenza o
in assenza di acqua, e vengono detti calori a umido e a secco. L'acqua accentua
l'azione del calore consentendo tempi più brevi e temperature più
basse del calore a secco. L'autoclave si basa sul calore a umido, con vapore a
15psi e con temperature che arrivano a 121°C , che in 20 minuti garantisce
sterilizzazione per piccoli volumi. La stufa, invece, si basa sul calore a
secco, utilizzando temperature a 160°C per almeno due ore. Altri metodi con
l'utilizzo di calore sono il flamberaggio e l'ebollizione. I metodi fisici di
sterilizzazione sono le radiazioni UV e ionizzanti. Le radiazioni UV hanno
lunghezze d'onda di 260nm esplicano la loro funzione sterilizzante agendo sugli
acidi nucleici e creando dimeri di timina; le radiazioni ionizzanti, che
comprende raggi X e ?, invece sono molto penetranti e esplicano la loro
funzione sterilizzante ionizzando atomi e molecole o creando radicali liberi.
La filtrazione è un processo che si utilizza su substrati liquidi che
non possono essere esposti ad alte temperature; questa tecnica utilizza filtri
con piccola porosità. I primi erano di porcellana (Chamberland) o di
terra ( Berkfeld) che avevano pori di diametro di pochi micron, oggi invece si
utilizzano filtri con pori da 50 Å. I metodi chimici di sterilizzazione
sono molto tossici anche per l'uomo, essi sono rappresentate da: l'ossido di
etilene, vaporizzazioni di formaldeide, glutaraldeide, propilattone, H2O2 e
infine ipoclorito. L'efficacia di uno sterilizzazione si fa valutando la sua
azione in date concentrazioni sulle spore, se efficace sulle spore è
efficace su tutti gli agenti. La disinfezione si fa con alcoli, con soluzioni
al 50%-70% che denaturano le proteine, oppure con alogeni del cloro, come
l'ipoclorito,di sodio al 5% o alcoli di iodio al 20%. I fenoli e i suoi
derivati anche a basse concentrazioni danneggiano le membrane e a alte fanno
precipitare le proteine; questi composti vengono usati nei saponi commerciali.
I saponi non sono battericidi ma ne facilitano la rimozione. La
pastorizzazione, ideata da Luis Pasteur, è un processo che viene
utilizzato nella conservazione dei cibi che non uccide le spore, cuocendoli a
62°C per 30 minuti (bassa) o a 72°C per 15 minuti(alta) consentendo di
conservare i gusti.
ESOTOSSINE
Le tossine batteriche sono veri e propri veleni, responsabili delle lesioni
anatomopatologiche e del quadro clinico consequenziale, rappresentano il vero
potere patogeno del batterio. Le tossine si dividono in esotossine e
endotossine: le esotossine vengono liberate, sebbene per modalità
diverse, dal batterio, e diffondibili nell'ospite; mentre le endotossine, che
sono rappresentate dall'LPS, non si diffondonose se non come risultato della
lisi del batterio. Altre differenze tra le due sono: che mentre le esotossine
sono polipeptidi, l'LPS è un lipopolisaccaride.; le esotossine sono
termolabii, mentre LPS è termostabile; infine mentre le tossine si
inattivano, diventando tossoidi (antitossine) e che conservano
l'antigenicità, e utilizzabili nei vaccini, l'LPS non può essere
inattivato, quindi non può essere utilizzato per fare dei vaccini.
I batteri che causano le malattie unicamente con tossine sono: Corynebacterium
diphtheriae che causa difterite, Bordetella pertussis che causa la pertosse,
Vibrio cholerae che causa il colera, Bacillus anthracis che causa il carbonchio,
Clostridium botulinum che causa il butolino, Clostridium tetani che causa il
tetano, Escherichia coli enteroemorragica che causa diarrea sanguinolenta e
sindrome uremica emolitica.
Una esotossina è formata da due subunità A e B, sintetizzate come
unica proteina e tagliata proteoliticamente e legate da ponti disolfuro. La
subunità A ha attività enzimatica, e costituisce la parte
tossica, che attraverso la subunità B, non tossica, capace di
riconoscere il suo sito d'azione della tossina e legarla alla membrane e
permettendo l'entrata o per endocitosi o per la formazione di un foro; la
subunità B è la parte antigenica, unavolta riconosciuta si separa
dalla subunità A. Alcune tossine sono codificate da plasmidi, che
possono essere trasferiti da un batterio ad un altro: la tossina di Vibrio
colerae può essere trasferito a Escherichia coli che diventa capace di
produrre tossine. A volte la capacità di produrre tossine non è
propria della specie ma del ceppo: alcuni ceppi di una specie sono tossigeni
mentre altri no ( Corynebacterium diphtheriae ). Le tossine agiscono in diversi
modi: danneggiando la membrana, o come i batteri piogeni formando pus, o
formando tramite tossine citolitiche ascessi, oppure inibendo proteine o la
loro sintesi bloccando il ribosomi a vari livelli, oppure attivando secondi
messaggeri e portando all'apoptosi. Alcune tossine attivano il sistema
immunitario più del normale provocando lo shock, uno di questi è
per esempio lo streptococcus aureus che replicandosi attiva tutti i linfociti,
in questi casi si dice che la tossina è un superantigene. Altre tossine
vengono dette neurotossine queste inibiscono l'attività dei
neurotrasmettitori.
Il tetano è causato da una tossina detta neurospasmina, questa tossina
riconosce dei recettori sui neuroni inibitori dei muscoli, e inibisce la loro
trasmissione. La neurospasmina ha come effetto primario la contrazione spastica
dei muscoli facciali in quello che viene chiamato risus sardonicus, poi
proandosi in tutto il corpo causa l'opistotono, ossia la contrazione
spastica di tutto il corpo, che portano alla morte per asfissia; questa tossina
è codificata da un plasmide.
La tossina difterica ha come suo recettore su un gran numero di tipi cellulari;
essa colpisce la sintesi proteica; le cellule morte formano psedomembrane che
spesso invadono l'arco palatino. La morte cellulare avviene perché la cellula
perde la capacità di produrre proteine agendo su l'ADPribosilazione del
elF-2 (fattore di elongazione); questo enzima ha il compito di spostare
l'amminoacido dal sito accettore a quello donatore. La azione del batterio
è spesso limitato al sito d'ingresso, ma la tossina provoca gravi danni
al cuore e al sistema nervoso;.
La tossina del bacillus anthracis, che ha come bersagli tutte le cellule,
è composta da tre subunità : il fattore edematoso I, che provoca
edemi se legato al fattore protettivo II, che a sua volta induce la sa di
anticorpi e conferisce immunità agli animali, e il fattore letale III
che causa la morte di piccoli animali se legato al fattore II. Il carbonchio,
causato della tossina, si manifesta con pustole maligne o come broncopolmonite,
che alla fine provocano la morte.
La tossina butolinica, prodotta da Clostridium butolinum, ha come bersaglio le
sinapsi motorie provocando una paralisi flaccida. Questa tossina può
essere di sette tipi, diversi solo dall'antigene non nell'effetto che portano,
i sette tipi sono: A , B , C , D , E ,F e G; le A, B, E e F sono quelle che
attaccano l'uomo. Alla fine la malattia causa la morte per asfissia, causata
dalla paralisi flaccida dei muscoli respiratori. Oggi vengono usati,
insensatamente, per l'eliminazione delle rughe.
I livelli in cui una esotossina agisce sono tre: a livello epiteliale, a
livello connettivale e vascolare, a livello di organi e tessuti. L'effetto di
una tossina può essere: letale, pirogeno, tossico sulla cute causare
eritemi o separare l'epidermide dal derma, o, infine, citolitico.
LA FLORA BATTERICA
In condizioni normali circa 200 specie batteriche diverse convivono con il nostro
corpo: sulla pelle crescono in gran quantità staphilococchi, le
neisserae proliferano nelle mucosa della faringe, le enterobatteracee vivono
normalmente nell'intestino; questa viene detta flora normale, diversa da
persona a persona. Molte interazioni tra batterio e ospite sono utili alla
sopravvivenza di quest'ultimo: alcuni enterobatteri producono la vitamina B, i
lactobacilli presenti nella mucosa vaginale la rendono altamente acida
proteggendola da batteri patogeni, alcuni batteri stimolano alla crescita di
tessuti come l'intestino o il sistema immunitario. Un esempio dell'importanza
della flora viena dal fatto che i bambini con flora non sviluppata se attaccati
da staphilococcus aureus producono la sindrome della pelle scottata, questo non
accade negli adulti con flora sviluppata. Nel concetto di flora è molto
importante il concetto di tropismo, ossia la capacità dei batteri di
poter vivere solo in alcuni tessuti che presentano le condizioni necessaria
alla vita; i batteri sui tessuti formano BIOFILM ossia uno strato complesso di
microrganismi contraddistinta dalla secrezione di una matrice adesiva e
protettiva.
INTERAZIONI BATTERIO OSPITE
La natura del rapporto tra ospite e batterio può essere di diverso
genere, che per quanto riguarda la flora normale è un rapporto
simbiotico, nel quale sia il batterio che l'ospite ne traggono vantaggio. Il
commensalismo è una condizione di completo indifferenza reciproca, nel
quale, sia il batterio che l'ospite utilizzano le risorse non compenetranti non
procurandosi ne danno ne vantaggio alcuno. Il parassitismo è un tipo di
associazione in cui il microrganismo vive a spese dell'ospite. Si definisce un
parassita opportunista un germe capace di svolgere attività patogena,
ossia di dare malattia solo in ospiti delibitati, mentre di solito è
saprofita, ossia non causa malattia (i trapiantati di valvole cardiache possono
essere attaccati da staphilococcus epidermidis, che solitamente è un
saprofita). Un parassita obbligato se è presente non può altro
che dare malattia; spesso però persone portatori di parassita obbligato,
non è affetto da malattia in questo caso si dice portatore sano.
La patogenicità è l'abilità del microrganismo di dare
malattia, tale abilità non è del tipo tutto o nulla, infatti
alcuni batteri danno malattia solo in alcuni tessuti non in altri (come
Eschelichia coli nelle vie urinarie). La patogenicità dipende dai
fattori di virulenza, ossia quei fattori come la sua stuttura o le tossine che
danno malattia.
La contaminazione è un contatto accidentale, e transitorio, tra
microrganismo e ospite, questa può essere interrotto e non produrre
nulla, o essere origine di successivi enenti. L'infezione è, la
moltiplicazione del germe, di solito su cute o le mucose, che non produce danni
oggettivabili, si dice che l'ospite è infetto, ed è capace di
trasmettere l'infezione. La malattia indica la moltiplicazione, ossia
l'infezione, con penetrazione, che causa danno anatomofunzionale più o
meno significativo. Le malattie sono classificabili in tre fasi: subclinica, in
cui non sono manifesti i sintomi; acuta , quando i sintomi sono manifesti che
dura meno di sei mesi; e cronica che dura più di sei mesi con sintomi
manifesti.
La virulenza è la capacità della misura di dare malattia in
condizioni naturali, essa è data da tre fattori: la capacità di
invadere l'ospite, la produzione di tossine, la resistenza alla terapia. La
capacità di invadere è data da l'adesione, che dipende dal
tropismo, e dalla penetrazione, cioè la capacità di produrre
enzimi che facilitano l'invasione di tessuti la neutralizzzazione delle difese
dell'ospite. La resistenza alla terapia è un nuovo fattore che è
so dal 1950.
Le infezioni sono: acute in cui la malattia dura poco e inoltre, la malattia
è autoremittente e autolimitante; acute con complicanze, in cui la
malattia procura disagi postumi; latenti, in cui dopo tempo dalla guarigione
ritorna con sintomi diversi; cronica persistente, che dura tempo e persiste;
cronica con sintomi tardivi, che dopo tempo vengono procurati disagi; e cronica
degenerativa,che si aggrava con il tempo, procurando la morte. Le infezioni
possono essere localizzate, se sono non si estendono a più di un punto,
o sistemica in cui raggiungono il sangue o la linfa e si disseminano in tutto
il corpo.
Le porte di entrata sono i distretti in cui comincia la colonizzazione, queste
possono essere artificiali o naturali: le prime avvengono con la
scominazione delle difese naturali da parte di agenti esterni, tramite danni
tessutali o per inoculazione nei capillari da parte di artropodi, queste
modalità di trasmissione vengono dette parenterali; le vie naturali sono
quelle per cui la trasmissione avviene attraverso il contatto con cavità
e tessuti, distinguiamo la trasmissione inalatoria, che avviene tramite il sistema
respiratorio, la trasmissione orofecale, che avviene tramite l'apparato
digerente, la trasmissione sessuale, che avviene tramite il tratto urogenitale,
e la trasmissione transplacentare. Affinché un germe sia patogeno deve
colonizzare il corpo: la prima fase viene detta adesione, che solitamente
avviene nella porta di entrata, grazie all'interazione recettoreantirecettore,
specifico da un germe patogeno a un altro, su u dato terreno, a questo punto
avviene l'invasione. L'invasione o penetrazione non è essenziale alla
patogenicità, i quanto i batteri che producono tossine hanno un'azione
patogena a distanza; gli esoenzimi sono i depositari del'invasività, che
agendo su vari substrati permettono l'invasione nei tessuti. La gonorrea è
causata da Neisseria gonorrhoeae (diplococco Gram negativo aerobio), che
tramite i pili aderisce alla mucosa vaginale, qui comincia la colonizzazione,
prolifera nella sottomucosa producendo pus, ma non entra in circolo, causando
un processo flagogistico. Il tifo è causato da Salmonella typhi
(enterobattero Gram negativo aerobio facoltativo), che attraverso cibi e acqua
inquinati da feci entra nell'intestino, invade l'ospite e entra in circolo,
causando febbri che arrivano anche a 40°C, enteroraggie, e peritoni.
ANTIBIOTICI E CHEMIOTERAPICI
Gli antibiotici sono farmaci naturali, tossici e specifici, ad altissima
selettività antibatterica, prodotti naturalmente da microrganismi. Gli
studi di Ehrlich, atte a trovare un proiettile magico, che si basavano sul
fatto che i coloranti di laboratorio coloravano selettivamente le cellule
batteriche, ma non le umane. Questi studi portarono alla scoperta del prontosil
(sulfamidico) e del salvarsan: il primo attivo contro gli streptococchi, il
secondo attivo contro Treponema pallidum, agente eziologico della sifilide. Nel
1929 Alehander Flaming scopre casualmente la penicillina, il primo vero
antibiotico, che nel 1949 entra in commercio; si calcola, che la scoperta degli
antibiotici, abbia allungato la prospettiva di vita di almeno 20 anni nei paesi
industrializzati.
La differenza tra un chemioterpico, come il prontosil, e un antibiotico, come
la penicillina, sta nel fatto che i primi sono composti di sintesi, i secondi
sono invece naturali; entrambi però hanno un azione specifica antibatterica.
Un batteriostatico è una sostanza che arresta la crescita di una
popolazione batterica, una volta eliminato, questa ricomincia; un battericida
è invece, una sostanza che uccide selettivamente i batteri. La
selettività dell'antibiotico o di un chemioterapico, e date dal fatto
che questi attaccano molecole che si trovano sul batterio, ma non sull'ospite.
Un chemioterapico o un antibiotico, agisco su determinate classi, queste classi
vengono dette spettro d'azione del antibiotico; ad esempio al penicillina
agisce solo sui Gram positivi, che sono dunque il suo spettro d'azione.
L'azione di un antibiotico è molto varia: alcuni inibiscono la sintesi
della parete come la penicillina o la cephalosporina, altri come la gentamicina
inibiscono la sintesi proteica, altri inibiscono la funzionalità della
membrana, altri ancora, la sintesi di acidi nucleici o sono inibitori
competitivi con i substrati.
La penicillina viene prodotta da Penicillina notatum, che in realtà
è composta di un insieme di sostanze con un anello beta-lattamico; oggi
vengono dette betalattamici di cui la penicillina è il precursore.
L'azione della penicillina è inibire la formazione del legame crociato
fra i tetrapeptidi della mureina, competendo con il legame fra il tetrapeptide
e la carbossipeptidasi. Lo spettro d'azione della penicillina sono i Gram
positivi, anche se alcuni betalattamici aggrediscono i Gram negativi come la
ampecillina.
Gli amminoglicosidi, il cui capostipite è la streptomicina, prodotta da
Strptomyces griseus, di cui oggi ne conosciamo molti altri come la amikomicina,
la gentamicina, la miomicina; gli amminoglicosidi sono inibitori della sintesi
proteica, si legano irreversibilmente all'unità 30S dei ribosomi non
permettono la traduzione del segnale. La spettro d'azione sono sia i Gram
positivi che i negativi, gli amminoglicosidi, sono stati i primi
antitubercolari.
I sulfamidici, la prima classe di chemioterapici, di cui fa parte il salvarsan,
sono composti da sulfanilamide, che sostituisce l'acido para amminobenzoico
precursore dell'acido folico utilizzato nella sintesi delle purine; esso
è un batteriostatico sia per i Gram positivi che i negativi.
L'uso continuo e spesso indiscriminato ha favorito e ha selezionato la nascita
di ceppi farmacoresistenti. La resistenza può essere inerente alla
specie, per la presenza di barriere o altro, come la membrane esterna dei Gram
negativi, oppure acquisita tramite mutazioni o per scambio di materiale
genetico, che avviene per trasformazione, ossia il processo tramite proteine di
batteri morti passano a batteri vivi, o per coniugazione. I meccanismi con cui
si attua la resistenza ai farmaci sono: sintesi di molecole che inattivano il
farmaco (betalattamasi), l'alterazione morfologica del sito di azione del
farmaco. È sa notare che la resistenza in Staphilococcus aureus, primo
batterio a essere curato con la penicillina, si è sviluppata nel 1950,
un anno dopo la messa in commercio della penicillina.
ISOLAMENTO E INDIVIDUAZIONE DI AGENTI INFETTIVI
La diagnosi di malattia è un difficile processo che comincia con la
raccolta di un campione, questo processo è importantissimo è deve
essere svolto nel giusto modo, e la successiva strisciatura del campione su di
un terreno solido. Il terreno solido sarà non selettivo se il campione
sarà monomicrobico, ossia deriva da terreni solitamente sterili come il
sangue, oppure sarà selettivo se il campione sarà polimicrobico,
ossia deriva da tessuti con una flora molto pronunciata come le feci. Il
terreno viene piastrato e incubato 24 ore per ottenere colonie pure; nei tre
giorni successivi si effettueranno test sulle colonie isolate: il primo giorno
si effettueranno test semplici, il secondo test con substrati, e il terzo test
sulla sensibilità ai farmaci (antibiogramma).
Il primo giorno si effettuano test semplici sulle colonie isolate con il fine
di fare una diagnosi presuntiva; per alcuni germi è possibile basarsi
sulla colorazione Gram, mentre per altri bisogna effettuare test come la
catalasi, che valuta lo sviluppo di ossigeno(positivo o negativo, strepto
staphilo) e l'ossidasi, che utilizzano la colorazione.
Il secondo giorno si effettuano test per una diagnosi i certezza, il cui scopo
è definire la specie del batterio che è causa della malattia, o
altre volte basta il genere. Le prove per una diagnosi di certezza utilizzano
substrati diversi onde vedere le reazioni del microrganismo, oppure si fanno
con terreni differenziali, in presenza di bile, o valutando la
sensibilità alla optochina, o osservando la produzione di H2S oppure con
prove sierologiche.
Il terzo giorno viene fatto l'antibiogramma, che serve per testare la
sensibilità di un agente batterico contro gli antibiotici e valutare una
terapia. Può essere fatto in due modi: con il metodo di Kirby-Bauer,
metodo qualitativo, e il metodo della determinazione della MIC, quantitativo.
Il metodo Kirby-Bauer si fa distribuendo uniformemente su un tampone una
colonia su di un terreno solido ricco di agar mullerito; su questo si applicano
dischetti imbevuti da antibiotici da testare si lascia a incubare 24 ore
lasciando crescere l'antibiotico e diffondere l'antibiotico; alla fine si
valuta l'alone di inibizione di crescita batterica. Il metodo Kirby-Bauer non
dice quanto farmaco usare, cosa che fa la MIC. MIC vuol dire minima
concentrazione inibente, e si fa con terreni liquidi, aggiungendo la stessa
quantità di batterio e aggiungendo quantità di antibiotico sempre
più diluite in diverse provette; si incuba 24 ore è si valuta la
minima concentrazione che ha inibito la crescita.
VIROLGIA
INTRODUZIONE
La parola virus viene dal latino e vuol dire veleno; esse sono entità
biologiche più piccole e semplici che esistono in natura, coevoluti con
i mammiferi, alcuni li infettano da sempre da sempre altri solo recentemente.
Nel1890 Ivanowski scoprì il virus del mosaico del tabacco, al tempo i
virus venivano classificati come filtrabili, in quanto, a differenza delle
tossine, filtravano attraverso filtri di 0.2µm; nel 1901 viene scoperto il
virus della febbre gialla, il primo virus umano. Solitamente la difficoltà
dello studio dei virus sta nel fatto che essi sono molto piccoli e
bisognò aspettare l'invenzione del microscopio elettronico perché
potessero essere studiati.
STRUTTURA DEI VIRUS
I virus sono organismi acellulari incapaci di replicarsi, questa loro
peculiarità li costringe a parassiti obbligati, ossia esplicano le
normali funzioni metaboliche o riproduttive se non a scapito della cellula
nella quale sono inseriti, non possiedono organuli, ne acido muramico ne
membrana esterna.
Le dimensioni vanno da 300nm(il più grande , il virus del vaiolo) a 20
nm, comunque molto più piccoli di staphylococcus aures che ha dimensioni
di 1000 nm. La loro struttura è molto semplice, sono cristalli proteici,
sempre di forma geometrica, e acidi nucleici, che possono essere a RNA o a DNA,
a doppio o singolo filamento, segmentato o unico filamento, a polarità
di messaggio o a antipolarità di messaggio (cioè possono essere
scritti nel verso in cui vengono sintetizzate le proteine o in quello inverso).
I virus sono insensibili agli antibiotici e ai chemioterapici, su di loro hanno
azione, invece, l'immunoprofilassi e gli antivirali.
Il virione è il corrispettivo della cellula per i virus, che è
formato da un involucro proteico che protegge il genoma all'interno dentro e
all'esterno della cellula, durante il trasporto da cellula a cellula.
L'ivolucro proteico deve essere stabile per non far degradere il genoma, ma
capace di liberarlo quando necessario, questa sua capacità viene detta
metastabilità.
Le caratteristiche comuni a tutti i virus sono: possiedono un acido nucleico
che ha tra i 3 ai 10-20 Kilobasi, un involucro proteico detto capside, e alcuni
possiedono un involucro lipidico detto pericapside o peplos, o envelope.
Il capside è la struttura proteica che ha il compito di proteggere il
genoma da enzimi litici e da condizioni avverse, inoltre nei virus privi di
pericapside svolge il ruolo di mediatore con i suoi antirecettore per
l'assorbimento. Il capside è composto da unità più piccole
dette capsomeri, visibili al microscopio elettronico come piccole strutture con
cavità centrale; a loro volta i capsomeri sono formati da protomeri a
loro volta formati da polipeptidi ripetuti, ciò è dovuto al fatto
che l'informazione genetica nei virus è molto limitata, quindi possono
fare solo pochi tipi polipeptidi. Il capside è una struttura
autoassemblante, come d'altronde le sue proteine,che sono in grado di
combinarsi con acidi nucleici, alcuni capsidi inoltre sono capaci di
riconoscere il nucleo della cellula ospite andare verso di questi. Le proteine
che compongono il capside, ai fini di creare strutture complete, si dispongono,
da poter formare il capside, in due simmetrie: l'elicoidale e l'icosaedrica.
La simmetria elicoidale è caratteristica dei virus animali a RNA eccetto
dei reovirus; i protomeri, tutti uguali, si dispongono a spirale, quindi il
capside è rappresentato da una struttura tubolare nella cui
cavità è presente il materiale gen.tico. La simmetria elicoidale
è tipica dei virus della rabbia, del morbillo e dell'influenza.
La simmetria icosaedrica, forma capsidi di forma solida a 12 vertici e a 20
facce triangolari (i più semplici), con un numero minimo di 3
subunità per faccia (con un totale di 60 unità) che nel caso del
poliovirus sono formate da tre protomeri VP1, VP2, VP3; i vertici sono formati
da cinque VP1. La formazione delle tre subunità avviene tagliando
proteoliticamente una proteina, codificata dal genoma, P1 in due
subunità 1ABC e 1D, la subunità 1ABC poi verrà tagliata in
1AB e in 1C; le tre subunita 1AB, 1C, e 1D legate costituiscono il capsomero.
Capsidi icosaedrici possono essere anche complessi, permettendo dimenzioni
maggiori, formati da 240 unità fatti di una stessa proteina detta esone
e di 12 unità ai vertici dette pentoni; con un totale di 252
unità; a volte come negli Adenovirus su ogni vertice si possono trovare
fibre proteiche con delle teste recettoriali.
L'involucro virale detto pericapside è di natura lipidica, essa origina
per gemmazione da una membrana dell'ospite, che può essere: quella
cellulare come per il virus dell'HIV, dal reticolo endoplasmatico come per il
virus vaccino, oppure dalla membrana nucleare come per l'erpesvirus. Oltre alla
membrana derivata per gemmazione, il pericapside contiene glicoproteine di
origine virale, che si assemblano in oligomeri con funzione recettoriali,
antigeniche, che permettono la fusione alle membrane e enzimatiche come le
neuramidasi; queste glicoproteine vengono sintetizzate usurpando le vie
sintetiche delle glicoproteine cellulari: il reticolo endoplasmatico sintetizza
proteine e N-glicosilazione, mentre l'apparato del golgi sintetizza
O-glicosilazione, che tramite vescicole secretorie saranno trasportate alla
membrana.
I virus più grandi possiedono altri componenti come: come la matrice che
ha il compito di mantenere il genoma all'interno del capside (Ortho- e
Para-myxovirus), il tegumento, una sostanza amorfa tra capside e
pericapside(Herpesvirus), enzimi virali, come RNA polimerasi-RNA dipendenti o
trascrittasi inversa, istoni (Papillomavirus) e proteine della cellula ospite
(Retrovirus).
TASSONOMIA ICTV E CLASSIFICAZIONE DI BALTIMORE
I virus si dividono in virus dei batteri, detti batteriofagi, e virus della
cellule eucaritiche , ossia virus di animali e piante. I virus come i batteri vengono
divisi in ordine, famiglie, generi e specie. L'ordine si indica con il suffisso
-virales (mononegovirales), nella pratica l'ordine è poco utile in
quanto non tutte le famiglie hanno un ordine. Le famiglie si indicano con il
suffisso -viridae (Paramyxoviridae), questa si basa sulla struttura, sulla
simmetria, sul rivestimento, sulle dimensioni, sul tipo di genoma; spesso il
nome è il risultato di questo criterio: Rabdoviridae, sono virus a
proiettile, i Picornaviridae sono invece piccoli virus a RNA. Il genere si
indica con il suffisso virus (Morbillivirus), i virus dello stesso genere sono
virus con caratteristiche comuni che li differenzia dei virus della stessa
famiglia ma di ordine diverso. La specie si definisce come una classe di virus
che costituiscono una linea replicativa e occupano una stessa nicchia
ecologica; cioè i virus che appartengono alla stessa specie devono
infettare lo stesso ospite e lo stesso tessuto, con l'utilizzo dello stesso
recettore, causando la stessa patologia, la stessa mappa genomica ma non
è detto la stessa sequenza nucleotidica. I tipi sono virus della stessa
specie che differiscono o per alcuni antigeni, e vengono detti sierotipi, o per
sequenza genomica, e vengono detti genotipi. Una classe politetica è un
gruppo di virus che hanno caratteristiche comuni nessuna essenziale. Il ceppo
non è una classificazione tassonomica, esso indica un isolato virale ben
definito appartenente alla stessa specie.
La classificazione di Baltimore viene fatta secondo quattro criteri e divide i
virus in dieci famiglie; i criteri utilizzati da Baltimore per la
classificazione sono: il tipo di genoma, il numero di catene, la
polarità, se è rivestito o no.
LO STUDIO DEI VIRUS
Lo studio della morfologia batterica avviene tramite:la microscopia
elettronica, il metodo più impiegato a partire dagli anni '50, che
tramite una risoluzione di 50-70 Å ci ha permesso di osservare le
strutture dei virus; l'immunoelettromicroscopia che permette di identificare e
visualizzare contemporaneamente il virus; cristallografia a raggi X, che
è possibile solo su virus cristallizzati e si basa sulla diffrazione dei
raggi X; e la risonanza magnetica nucleare su virus in soluzione. Lo studio al
miroscopio viene fatto tramite colorazione negativa fatta con soluzioni purificate
sali di metalli pesanti e di virus; oppure con colorazione positiva in cui si
utilizza la sensibilità di alcune componenti virali per alcuni sali:
uralinacetato di sodio per gli acidi nucleici.
LA REPLICAZIONE DEI VIRUS
Non tutte le cellule permettono la produzione di virus, quelle che lo
permettono, vengono dette permissive, mentre quelle che non permettono la
produzione di virus, ma permettono l'infezione vengono dette suscettibili. La
replicazione virale è un complesso processo a tappe, che comincia con
l'adesione o assorbimento.
I virus come i batteri hanno un tropismo, possono attaccare solo date cellule,
questa loro capacità è visibile durante l'adesione, che è
consentita dall'interazione tra gruppi chimici presenti sulla cellula, detti
recettori e dei gruppi ripetute molte volte sulla superficie esterna del virus
detti antirecettori virali, costituite da proteine del capside oppure
glicoproteine del pericapside. L'interazione tra il recettore e l'antirecettore
è un meccanismo del tipo chiave e serratura, in cui il antirecettore
riconosce solo un tipo di recettore, questa interazione può essere di
due tipi: a zip e a gancio. Il meccanismo a zip prevede che una volta
riconosciuto il recettore, l'antirecettore si leghi a questo favorendo la
penetrazione del virus tramite modificazioni conformazionali della membrana (
HIV, influenza); il meccanismo a gancio invece una volta legato non ne media
l'entrata (Poliovirus).
La penetrazione è la tappa seguente all'adesione, questo non è un
processo unico, ma una serie di processi, specifico per ogni virus, che hanno
in comune il fatto che necessitano di energia, energia che, la cellula ospite
utilizza per farla per permettere la penetrazione. I due meccanismi più
usati sono l'endocitosi e la fusione; il primo, l'endocitosi comincia con
l'adesione, il virus viene convogliato nella bristle coated pits, piccole
invaginazioni della membrana cellulare rivestite di clastrina, e qui
fagocitato, una volta entrato il fagosoma viene circondato da lisosomi,che fondono
il capside alla membrana e liberano il genoma; il secondo meccanismo la fusione
è possibile solo ai virus rivestiti: il pericapside virale si fonde con
la membrana, riversando al di dentro della membrana il nucleocapside. Il
meccanismo della fusione, che può essere o no pH dipendente, necessita
una proteina dette F che si trova sul pericapsde e favorisce la fusione e lo
scapsidamento.
La liberazione del genoma, o scapsidamento, è la terza tappa, spesso
contemporanea alla penetrazione, che comporta la liberazione del semplice
genoma o del nucleocapside. Anche questo processo differisce molto da virus a
virus, comunque per la maggior parte dei casi avviene tramite la proteina F
(Paramyxovirus) o dei lisosomi(Orthomyxovirus), oppure iniettando il genoma
tramite un poro(Picornavirus).
La traslocazione è un processo in cui il genoma arriva al nucleo,
infatti, per i i retrovirus e i virus e i virus a DNA, la replicazione avviene
nel nucleo, ma il semplice acido nucleico non sa arrivarci da solo, necessita di
sequenze cariofiliche presenti sul capside. Queste catene cariofiliche sono
sequenze amminoacidiche che riconoscono i pori nucleari, permetteno il
trasferimento del nucleocapsde fino dove avverrà lo scapsidamento, a
questo punto comincerà la replicazione.
I virus a RNA stampano il loro RNA da stampi a RNA, solo i retrovirus
necessitano intermedi a DNA. I virus a RNA pol+ (con polarità di
messaggio, poliovirus con DNA infetto) possono direttamente andare sui ribosomi
a essere tradotti in proteine giganti che verranno tagliate in proteine
più piccole. La copia del RNA, avviene prima scrivendo molte copie a RNA
pol-, che serviranno da stampo per RNA pol+, che a loro volta andranno a
tradurre in proteine e enzimi. I virus a RNA pol- (con antipolarità di
messaggio)devono essere prima tramite trascrittasi inversa, già presente
nel virus, in copie a RNA pol+, che codificherà per proteine e altre
copia di RNApol-. I virus a DNA, si replicano nel nucleo, questo solo dopo aver
tradotto proteine precoci, solitamente enzimi, dopo la replicazione produce
proteine tardive, strutturali.
L'incorporazione del genoma è un processo in cui il genoma e le proteine
strutturali producono nuovi virioni, tale riconoscimento avviene nella sede di
replicazione, e prevale con l'aumento delle proteine e del genoma. Il
meccanismo avviene grazie al riconoscimento del genoma da parte di una proteina
virale come avviene per i retrovirus, in cui la proteina nP riconosce il RNA, e
poi con il capside forma il virione.
A questo punto avviene il rilascio i virioni maturi per lisidella cellula, o
per gemmazione, e in questo caso si produce il pericapside; alcuni virus come
gli Herpesvirus, prima gemmano dalla membrana nucleare, poi lisano la cellula
INTERAZIONI FRA VIRUS E OSPITE
La patogenicità virale si svolge attraverso un effetto diretto e uno
indiretto. L'effetto diretto è dato dall'effetto citoplasmatico della
replicazione virale, e solo per alcuni virus dall'oncogenicità;
l'effetto indiretto è data dalla risposta dell'ospite all'infiammazione,
ossia dall'immunopatogenesi, che è la patogenicità portata dal
sistema immunitario.
L'effetto diretto è dato soprattutto dall'effetto citopatico(ECP), ossia
l'effetto che ha il virus quando si replica nell'ospite, causato da alterazioni
morfologiche della cellula visibili al microscopio elettronico. Gli effetti
citopatici cambiano da cellula a cellula, da virus a virus; l'azione dannosa
è esplicata con alterazioni della forma come il ballonig, in cui la
cellula da piatta diventa sferoidale; provocando la lisi cellulare (sul vetrino
si osservano placche litiche); oppure creando per fusione scincizi, ossia
cellule con molti nuclei; provocando permeabilità alterate delle
membrane; oppure con l'accumulo di componenti virali nella cellula; apoptosi o
vacuolizzazione(zone più chiare). Non tutti i virus danno l'effetto
citopatico.
Effetto iceberg si definisce come l'effetto per cui benché molte persone
vengono incontro a virus o batteri, ma molte mene si ammaleranno e ancora meno
si ammaleranno, e ancora meno moriranno. Un esempio a ciò ci viene dato
dal poliovirus, che entra nell'intestino, comincia a replicarsi per raggiungere
il sangue, va al sistema nervoso sove si replicherà causando la
polimelite: a ognuno di queste tappe il virus può essere fermato e non
portare alla malattia.
La persistenza è la caratteristica dei virus secondo la quale anche dopo
la guarigione il virus può rimanere; questa caratteristica è
dovuta al fatto che la risposta può essere elusa o essere inefficace; il
virus può vivere uno stato di latenza dal quale si può
risvegliare oppure resistere sotto forma di DNA integrato(lisogenia, con la
repressione di geni e integrazione) o con espressione repressa (andando
incontro a cicli abortivi, producendo proteine ma non virioni completi). La
persistenza può essere attuata grazie a variazioni antigeniche, o
producendo sostanze che inibiscono le difese oppure passando per vie non
ematiche come gli assoni.
La trasformazione è un processo a tappe che colpisce le cellule ospite,
in cui: dapprima la fa moltiplicare all'infinito, poi perduto l'inibizione da
contatto, subisce mutazioni che comportano a una forma diversa e a alterazioni
metaboliche; la trasformazione portano alla formazione di tumori. La
trasformazione è dovuta a: presenza di geni tumorigenici
(oncogèni) nel genoma virale infettante (SV40), cis-attivazione
dell'espressioni di oncogèni cellulari (EBV) per integrazione del genoma
virale, trans-attivazione tramite proteine virali, generazione di stati
infiammatori cronici che portano alla trasformazione cellulare sul lungo
periodo. I virus che portano alla trasformazione vengono detti oncovirus.
LA DIFESA IMMUNITARIA
Le difese contro le infezioni sono data da una immunità aspecifica, e
una specifica; il primo è un meccanismo non specifico e innato, sempre
uguale a se stesso, il secondo invece è specifico per il suo agente
scatenante. Fanno parte del primo le cellule dendritiche, i macrofagi, le
cellule natural killer, i mediatori dell'infiammazione e quelli del
complemento; del secondo tipo fanno parte i linfociti B e i linfociti T, che si
attivano solo dall'agente di cui hanno memoria di una precedente infezione, non
permettendone una seconda. L'immunità specifica, che si può
definire come acquisita, può essere naturale attiva, causata da
infezione da patogeni che dura tutta la vita, o essere naturale passiva, che
provenire dalla madre, e durare poche settimane; infine esiste
un'immunità acquisita può essere artificiale attiva, prodotta dai
propri linfociti, acquisita tramite la vaccinazione che dura da anni o in
alcuni casi per sempre, oppure essere artificiale passiva tramite la
sieroprofilassi, che dura massimo dei mesi (influenza con IgA).
I linfociti B e T fanno parte del sistema immunitario, hanno pari peso al
cervello o al fegato, li possiamo dunque considerare un organo circolante
composto da 2*1012 cellule. I linfociti B e T sono il 25% delle cellule del
sangue e il 100% della linfa. I linfociti B sono responsabili della risposta
umorale, essi producono immunoglobuline, anticorpi in siero che non sono
presenti in condizioni normali. I linfociti T sono coinvolti nella risposta
mediata da cellule, in quanto tramite citochine e linfochine coordinano
l'azione delle altre cellule. I linfociti B e T girano tutto il corpo nell'arco
di 24 ore.
Il siero è la parte liquida del sangue dopo la coagulazione e la
centrifugazione, se contiene però anticorpi contro un antigene
specifico, viene detto antisiro; i rapporti tra fra antigeni e anticorpi,
vengono studiati dalla sierologia, essendo gli anticorpi(o gamma
immunoglobuline o immunoglobuline) la frazione di proteine seriche, prodotte
dai linfociti B naturalmente, addette al legame con antigeni (per lo più
microbici).
La forma di un anticorpo è a Y dove il corpo viene detto catena pesante
(due per monomero), mentre i bracci vendono detti catene leggere (due per
monomero), sia le catene leggere che le pesanti possiedono una parte fissa,
sempre uguale, e una parte specifica che si lega all'antigene; ponti di solfuro
permettono alle catene pesanti e alle leggere di restare uniti. Gli anticorpi
sono il 20% delle proteine plasmatiche, e si dividono in cinque classi:
G,M,A,DeE, tutte diverse per funzione e struttura.
Le IgG sono le più efficaci, e sono l'80% delle proteine sieriche, e
sono costituite da monomeri; esse sono le uniche che possono essere trasmesse
al feto, e il loro compito è opsonizzare , ossia preparare alla
fagocitosi da parte del macrofago, neutralizzano l'antirecettore, impedendo che
si leghi al recettore e infine proteggono il feto.
Le IgM sono le prime a formarsi, ma sono le meno efficaci; si formano dopo
7-l0giorni dopo l'infezione primaria e hanno un picco verso 2-3 settimane,
sendo in tre mesi. Sono il 5%-l0% delle proteine sieriche, e sono
costituite da pentameri uniti con una joing chain.
Le IgA sono dimeri tenuti insieme da catene J, e rappresentano il 10%-l5% delle
proteine sieriche,vengono riversate su epiteli e mucose, tramite transcitosi, e
impediscono l'insorgere di infezioni, hanno un emivita di 6-7 settimane.
La risposta primaria viene fatta dalle IgM, dura poche settimane o mesi, e
raggiunge il suo picco di concentrazione dopo7-20 giorni, essa ha poca
affinità con l'antigene. La risposta secondaria, più lenta, viene
fatta dalle IgG e dalle IgA, il cui effetto può durare molti anni,
inoltre ha un alta affinità con l'antigene. Le conseguenze portate dalla
risposta anticorpale, sono, l'agglutinazione (vengono tenute unite),
l'opsonizzazione, la neutralizzazione, l'attivazione dell'infiammazione e del
complemento.
I VACCINI
I vaccini sono preparazioni che contengono un agente patogeno modificato, che,
una volta inoculato sia capace di indurre una risposta immune, protettiva
contro il patogeno. I vaccini hanno uno scopo puramente preventivo,la cui
efficacia è dovuta dal fatto che acquisiamo l'immunità
attivamente. Per la preparazione dei vaccini si utilizzano agenti vivi
attenuati tramite trattamenti fisici o chimici (antipolio di Salk, inattivato
con formolo), o attraverso culture successive in condizioni diverse dalla
normali (antipolio di Sabin, fatto da mutanti), questi attivano la risposta ma
hanno perso il loro potere patogeno. Altri vaccini vengono purificati, ossia,
vengono inoculati porzioni di agente patogeno, come polisaccaridi del peplos o
strutture del capside (stesso meccanismo con i tossoidi delle tossine:
antitetanica, antidifteritica). Altri ancora sono vaccini ottenuti con
l'ingegneria genetica, tramite i lieviti (antiHBV). Infine se molti persone
appartenenti di una popolazione sono vaccinati per una malattia, sui non
vaccinati si istaura un'immunità di branco: se molti sono vaccinati sono
ridotti il numero di persone che le può trasmettere la malattia.
L'INFIAMMAZIONE
L'infiammazione è un processo in risposta a un danno a tessuti vascolarizzati
con lo scopo di recapitare materiali difensivi nella sede del danno. Questo
processo è attivato da tutti gli agenti capaci di provocare un danno e
si definisce come difesa aspecifica, in quanto qualsiasi sia l'agente il
processo è lo stesso. Possiamo distinguere una infiammazione acuta che
dura meno di sei mesi e di natura vascolare, e una infiammazione cronica che
dura molto di più, e inoltre di natura tissutale. Il primo avvenimento
di un'infiammazione è la dilatazione dei vasi e l'aumento della permeabilità,
che permette ai leucociti di migrare nell'essudato. La funzione dell'essudato
è facilitare la fagocitosi e la creazione di barriere atte a fermare
l'infiammazione. Quando è in atto un processo infiammatorio il vaso
diventa più appiccicoso e tramite un processo detto diapedesi i
leucociti possono uscire dai vasi, e per chemiotassi raggiungere il punto
dell'infiammazione. La fagocitazione avviene ad opera dei neutrofili, che
distruggono l'agente attraverso due meccanismi: uno tramite enzimi lisosomiali,
e un meccanismo ossigeno dipendente. Il primo distrugge il fagosoma fondendolo
al lisosoma distruggendolo; il meccanismo ossigeno dipendente invece tramite la
formazione di H2O2 e ipoclorito creano radicali liberi nel agente patogeno, che
lo distrugge. Le cellule coinvolte nell'infiammazione sono i linfociti T che
tramite le linfochine e le citochine attivano i macrofagi e i linfociti B che
producono gli anticorpi.
LA DIAGNOSI VIROLOGICA
La diagnosi immunologia d'infezione virale ha lo scopo di stabilire terapie
appropriate, se possibili, o studiare procedure preventive ai fini
epidemiologici. Ai fini della diagnosi immunologia distinguiamo due metodi: la
diagnosi diretta e la diagnosi indiretta.
La diagnosi diretta avviene con la ricerca di proteine o altri componenti
dell'antigene; ai fini diagnostici esistono varie metodiche dirette che
richiedono prima l'isolamento e l'identificazione con anticorpi:la titolazione,
la PCR, emagglutinazione (or HAI), ELISA diretto, identificazione con anticorpi
(immunofluorescenza) e tramite la microscopia elettronica.
L'emoagglutinazione sfrutta la capacità d'alcuni virus a concentrazione
elevate di riconoscere e di legarsi agli eritrociti detta emoagglutinazione, e
di formare numerosi ponti che portano all'agglutitinazione degli eritrociti. La
titolazione attraverso l'emoagglutinazione è fatta mediante
predisposizione in pozzetti seriali diluizioni d'agente virale e con l'aggiunta
di un volume prestabilito di sangue, e si calcola la diluizione minima capace di
dare emoagglutinazione, e si assegna a questa 1 UE, con questa si risale alla
concentrazione originale.
Il metodo ELISA diretto utilizza sospensioni solide d'anticorpi che hanno
specificità nota per l'antigene, a cui è aggiunto il campione.
L'agente virale si attacca all'anticorpo, e dopo aver rimosso con dei lavaggi
tutto ciò che non si è legato, si aggiunge un anticorpo con
specificità uguale a prima marcato con un enzima, attraverso lavaggi si
leva l'anticorpo non legato. L'attività enzimatica è connessa con
la quantità di virus.
L'immunoflurescenza è una metodica che utilizza anticorpi con
specificità per l'agente virale, marcati con sostanze fluorescenti, che
si attaccano ai virus; dopo il lavaggio siamo in grado di visualizzare con
lampade apposite i virus.
Le metodiche indirette o sierologiche, avvengono con la ricerca d'anticorpi
specifici per il virus che stiamo studiando. Le metodiche più usate
sono: la neutralizzazione, la fissazione del complemento, l'inibizione
dell'emagglutinazione e l'ELISA indiretto.
Le reazioni di neutralizzazione si basano sul fatto che anticorpi solubili,
fissatesi su antigeni molecolari o corpuscolari ne inibiscono l'attività
specifica, in questo caso siamo capaci di dire la minima concentrazione di
siero, ottenuta con diluizioni successive, capace di inibire l'attività.
L'ELISA indiretto si fa introducendo il siero in esame in una provetta dove
è stato fissato l'antigene in esame, in seguito si aggiunge antiIgG
umano fissato con un enzima e dopo il lavaggio si aggiunge il substrato;
l'attività enzimatica indica la concentrazione d'anticorpo nel siero.